Náplň podkapitoly:
1. Úvod do vyšších funkcí centrální nervové soustavy
2. Funkce specifických korových oblastí
3. Řeč
4. Paměť
_
Úvod do vyšších funkcí centrální nervové soustavy
Otázky, které vědce a filozofy již celé generace na neurovědách přitahují (tedy: „Co nás dělá tím, čím jsme? Co je naše vědomí? Jak vědomí vzniká?”), jsou právě ty, na než je nejtěžší nalézt odpověď a o kterých toho paradoxně víme nejméně. Kůra mozku je jednou z jeho nejhůře prozkoumaných částí. Většina našeho poznání pochází z epizodických kazuistik pacientů, jimž byla kůra buď při nehodě nebo během operace poškozena, popřípadě z experimentů s elektrostimulací různých korových oblastí.
Funkční morfologie kůry
Funkční část kortexu tvoří asi 100 miliard neuronů, které se nacházejí ve vrstvě o šířce pouhých 2 až 5 mm na ploše asi 0,25 m2. Ty jsou uspořádány do šesti korových vrstev:
1) Lamina zonalis
2) Lamina granularis externa
3) Lamina pyramidalis externa
4) Lamina granularis interna
5) Lamina pyramidalis interna
6) Lamina multiformis
Lamina zonalis
Vrstva na povrchu kůry. Obsahuje zejména dendrity pyramidových neuronů z vrstev III a V, dále také projekce z intralaminárních jader thalamu. Kromě toho je zde i malé množství Cajal-Retziusových neuronů, jež mají excitační vliv na pyramidové buňky.
Lamina granularis externa
Zde nalezneme množství kosíčkových buněk, které svou aktivitou inhibují pyramidové buňky III. vrstvy. Volně jsou zde i rozptýlené malé excitační pyramidové buňky.
Lamina pyramidalis externa
Obsahuje středně velké pyramidové neurony, které přijímají informace hlavně z hvězdicových interneuronů IV. vrstvy a thalamokortikální projekce. Vysílají komisurální vlákna do kontralaterální hemisféry a asociační vlákna do nejrůznějších oblastí ipsilaterální kůry.
Z lamina granularis externa (II.) a pyramidalis externa (III.) pochází většina korových asociačních a komisurálních drah.
Lamina granularis interna
Obsahuje interneurony různých tvarů, ale převažují hvězdicové neurony excitačního charakteru. Tato vrstva je velmi významná, neboť zde končí většina vláken thalamokortikální projekce (je jejím hlavním cílem). Mimo to rozvádí signály z thalamu do dalších vrstev. Zvlášť mohutná je v senzorických oblastech.
Lamina pyramidalis interna
Velké neurony této vrstvy dávají vznik projekčním vláknům do podkorových struktur. V motorických oblastech se v lamina pyramidalis interna nacházejí velké pyramidové neurony zvané Becovy buňky, jejichž vlákna jsou součástí pyramidové dráhy.
Lamina multiformis
Jak již název napovídá, obsahuje neurony různých tvarů. Objevují se zde jak interneurony, které ovlivňují činnost vyšších vrstev, tak buňky, jejichž axony dávají vznik kortikothalamické dráze – integrální součásti tzv. thalamo-kortikálního oscilátoru, o němž se soudí, že se podílí na udržení vědomí.
Vztah thalamu a kůry je velmi významný a obzvlášť patrný při přerušení projekčních vláken z thalamokortikální a kortikothalamické dráhy. Různá thalamická jádra nebo jejich funkční skupiny mají propojení s určitou oblastí kůry, se kterou funkčně souvisí. Vzájemně spolu velmi intenzivně komunikují. V případě přerušení této komunikace (vzájemných projekcí) je naprosto ztracena funkce odpovídající části kůry.
Lamina pyramidalis interna (V.) a lamina multiformis (VI.) jsou hlavním zdrojem projekčních vláken kůry. Lamina V vysílá projekce do striata, tekta, retikulární formace, mesencefala, prodloužené míchy a hřbetní míchy. Lamina VI do thalamu.
_
Funkce specifických korových oblastí
Už od dob antiky byly různým částem kůry přiřazovány rozdílné funkce. Těchto starých domněnek se chopil Korbinian Brodmann, který provedl pečlivý histologický rozbor kůry mozku a opravdu objevil 52 morfologicky rozdílných oblastí. Na základě tohoto objevu předpokládal mezi cytoarchitektonicky různými oblastmi funkční rozdíly.
Jakkoli je Brodmannova práce dnes již moderní technologií do jisté míry překonána, dala vznik široce užívané terminologii, která se i přes mnohé nedostatky stále respektuje. Zčásti proto, že nástupnické práce jsou nepoměrně složitější, a zčásti pro svou dlouhodobou zažitost a všeobecnou srozumitelnost.
Zajímavost:
Rozdělení kůry na funkčně specifické oblasti je evolučně relativně mladý fenomén. Někteří plazi – např. želvy – mají tzv. generalizovaný kortex, který vykazuje jednotnou morfologickou strukturu i funkci.
Senzorické a motorické oblasti jsou probrány v příslušných kapitolách. V této části textu se budeme zabývat asociačními oblastmi kůry.
Asociační oblasti
Asociační oblasti získávají a simultánně zpracovávají informace z mnoha senzorických i motorických oblastí stejně jako z podkorových struktur. Poněvadž právě tyto oblasti integrují a interpretují jednotlivé informace, vytvářejí tak celistvý „obraz reality”. Významnými asociačními oblastmi jsou:
1) Parieto-occipitální asociační oblast
2) Prefrontální asociační oblast
3) Limbická asociační oblast (diskutována v kapitole Limbický systém)
1) Parieto-occipitální asociační oblast
Tato oblast se nachází mezi somatosenzorickými oblastmi (rostrálně), vizuálním kortexem (kaudálně) a sluchovou kůrou (ventrálně). Získává informace ze senzorických oblastí, které ji obklopují. Tyto informace integruje a interpretuje. U člověka se dále rozčleňuje na podoblasti plnící vysoce specializované funkce.
Analýza polohy těla a okolních objektů
Podoblast v zadní parietální a horní occipitální kůře. Integruje informace zejména z vizuálního kortexu a somatosenzorické oblasti. Z těch tvoří vnitřní reprezentaci polohy, a to jak těla vůči okolí, tak prostorových vztahů mezi jednotlivými předměty.
Wernickeho oblast
Podoblast ležící kaudálně od primárního sluchového kortexu, v zadní části g. temporalis superior. Je obzvláště vyvinutá v dominantní hemisféře – u většiny osob v levé. Má zcela zásadní význam pro porozumění řeči, o níž budeme podrobně diskutovat dále v této kapitole.
Navíc zde sídlí celá řada intelektuálních funkcí. Wernickeho oblast je tak významná, že je někdy nazývána hlavní interpretační oblastí. Přicházejí do ní informace z většiny senzorických a asociačních areí, které integruje a zpracovává.
Při poškození této oblasti – například při cévní mozkové příhodě v levostranném povodí a. cerebri media – dochází k těžké poruše řeči: senzorické afázii. Postižený je při tomto typu afázie schopen rozeznat zvuky jednotlivých slov, ale již nedokáže pochopit jejich význam nebo utvořit významově koherentní větu. Stejně tak rozezná zápis slov, nechápe však jejich význam.
Ztráta celé oblasti učiní z postiženého jen „stín původní lidské bytosti” neschopný jakékoliv komunikace, nechápající jakýkoliv povel, zcela odkázaný na pomoc jiných osob. Nejedná se pouze o zevní projevy – ztrátu schopnosti komunikovat a jakoukoliv komunikaci chápat. Jde převážně o ztrátu většiny intelektuálních schopností.
Stimulace Wernickeho oblasti vyvolává celou řadu velmi zajímavých fenoménů, obvykle se jedná o velmi komplexní myšlenku. Stimulovaná osoba je totiž schopna si například vybavit celý proslov, který někdy slyšela, dávnou vzpomínku se všemi detaily včetně vůní a zvuků nebo třeba začne halucinovat.
Obecně lze říci, že Wernickeho oblast interpretuje motivy ve vnitřní reprezentaci reality, jež byly získány na podkladě senzorických vstupů.
Wernickeho oblast výrazně zasahuje i do procesu učení. Většina senzorických vstupů je před zapamatováním převedena do řečového ekvivalentu. Obvykle si pamatujeme stranu textu nikoli jako fotografii, kterou si vybavíme, ale jako verbální informaci. Tento jev má pravděpodobně souvislost s tím, že v útlém dětství získáváme většinu informací mluveným slovem. Až později si osvojíme dovednost číst a do procesu učení zapojíme výrazněji i psané slovo a s ním gyrus angularis.
Gyrus angularis
Nachází se v anterolaterální části occipitálního laloku. Často je považován za „vizuální asociační oblast” díky svému významnému propojení s vizuálním kortexem. Tato oblast rozeznává psaná slova zachycená vizuálním kortexem a informace přeposílá do Wernickeho oblasti, kde je dešifrován jejich význam. V každém okamžiku, kdy čtete tento text, probíhá masivní komunikace po ose vizuální kortex → g. angularis → Wernickeho oblast.
Při poškození g. angularis není postižená osoba schopná rozeznat psané slovo, přestože ostatní jazykové funkce jsou naprosto intaktní. Takovéto oblenění funkce g. angularis nabývá různé intenzity a nazývá se dyslexie.
Pojmenování objektů
Podoblast ventrálně v rostrální části occipitálního laloku. Integruje informace o názvu předmětu s vnitřní reprezentací daného objektu, která pochází z vizuálního kortexu. Nejdříve ale musí být název rozpoznaného objektu osvojen prostřednictvím vstupu ze sluchové kůry a jeho vnější vzhled z kůry zrakové.
Jednoduše řečeno: Pokud jste se dříve naučili, jak se určitý předmět nazývá (např. vám to někdo řekl), a vy jej znovu uvidíte, je k němu díky této podoblasti automaticky přiřazen název.
2) Prefrontální asociační oblast
Oblast rostrálně před motorickými oblastmi, se kterými extenzivně spolupracuje na plánování komplexních pohybových motivů a sekvencí. Aby byla schopna efektivně se účastnit plánování motorické aktivity, je propojena masivním subkortikálním svazkem asociačních vláken s parieto-occipitální oblastí, od níž získává přesnou reprezentaci polohy těla a okolních předmětů. Plán pohybu z prefrontální oblasti pak prochází okruhy bazálních ganglií.
Prefrontální asociační oblast je také zcela esenciální pro myšlení a vznik reprezentace „vnitřního hlasu”. Ten zde vzniká pravděpodobně kvůli úzkému propojení s motorickými oblastmi včetně mluvidel.
Tato oblast pravděpodobně také zajišťuje funkce pracovní paměti, která dočasně uchovává nové vědomé informace z ostatních senzorických oblastí. Díky pracovní paměti dokážeme plánovat budoucnost, řešit složité matematické a filosofické problémy, kontrolovat své chování (porovnávat plánovaný vzorec chování s morálními zásadami) aj. Všechny tyto schopnosti ztrácíme při poškození prefrontální kůry. V minulosti šlo o časté poškození v důsledku prefrontální lobotomie, nadužívaného zásahu u psychotických pacientů. Prefrontální lobotomie spočívala v přerušení propojení prefrontálního kortexu a dalších částí mozku.
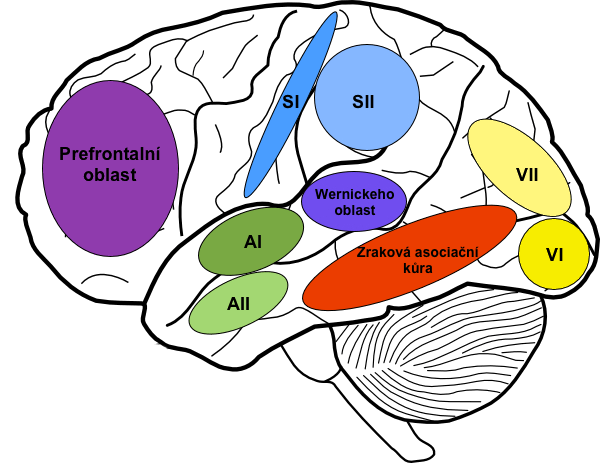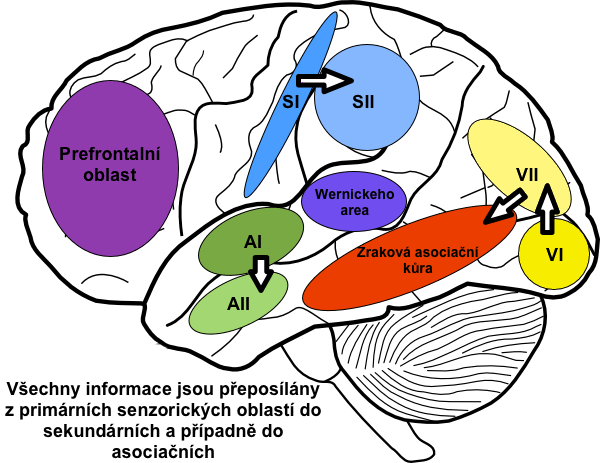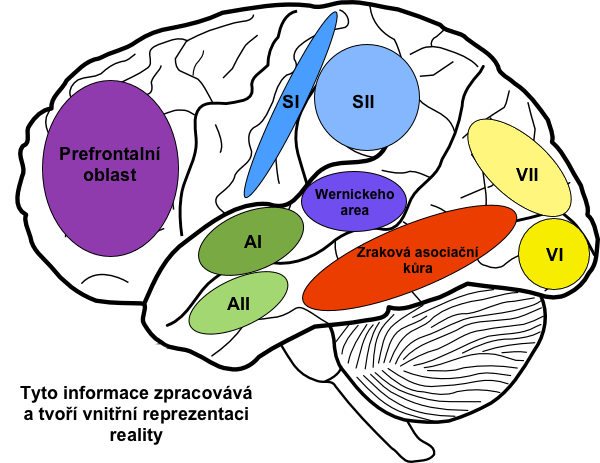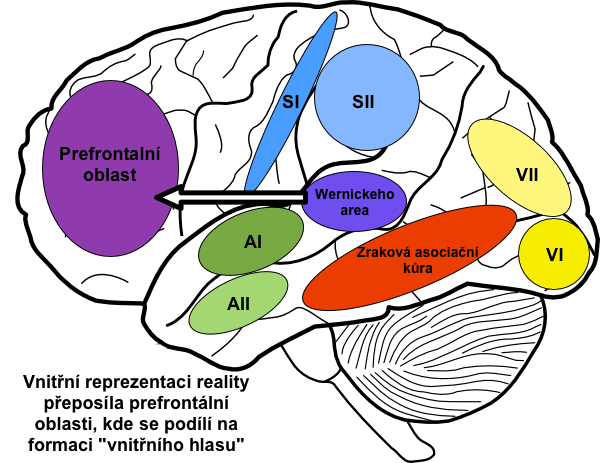
Brocova oblast
Brocova oblast (případně Brocovo centrum) se nachází zčásti v prefrontální asociační kůře a zčásti v premotorické oblasti. Odpovídá za formaci slov pomocí synchronizace pohybů dýchacích svalů se svaly laryngu a úst. Zde také vzniká plán a je iniciována artikulace slov a krátkých frází.
Oblast pro rozpoznání tváří
Je to vlastní samostatná asociační oblast. Pravděpodobně se tato část kůry vyčlenila v průběhu evoluce, když se předci současných lidí začali výrazněji socializovat. Nachází se na mediálním povrchu části occipitálních a temporálních laloků. Má četná propojení s vizuální kůrou a limbickým systémem. Byla objevena v souvislosti s prosopagnosií, postižením, při kterém není pacient schopen rozeznat tváře. Prosopagnosie je jediná patologie, jež vznikne při destrukci tohoto relativně velkého okrsku kůry.
Lateralizace
Pojem lateralizace znamená, že jedna hemisféra je dominantní. Konkrétně ta, ve které je více vyvinuta Wernickeho interpretační oblast, g. angularis, další řečová centra a motorické oblasti.
U 95 % osob se jedná o levou hemisféru, a to jak u pravorukých, tak levorukých jedinců. Tento jev se dá pozorovat již při porodu, kdy většina novorozenců vykazuje o 50 % větší Wernickeho oblast vlevo. Zajímavé je, že pokud je novorozenci tato oblast poškozena, adaptuje se na dominantní úlohu pravá hemisféra.
Lateralizace se vyskytuje u většiny savců. Teorie jejího vzniku je taková, že během procesu učení v raném dětství je přirozeně více užívána ta hemisféra, která měla v perinatálním období více vyvinutý temporální lalok (respektive jeho Wernickeho oblast). V průběhu učení je pak více zapojována do interpretace informací i exekuce pohybů. Prodělává tak více změn z hlediska neuronální plasticity.
U zbylých 5 % osob se obvykle vyvinou obě hemisféry bez lateralizace. Pouze velmi vzácně je dominantní pravá hemisféra.
Brocova oblast je také více vyvinuta na dominantní straně. Vzhledem k tomu, že odpovídá za formaci slov pomocí synchronizace pohybů dýchacích svalů se svaly laryngu a úst, jakýkoliv zásah do přirozeného vývoje lateralizace může vést k poruše řeči. Například pokud dítě píše levou rukou a je násilně přeučováno, začne často vykazovat koktavost, přestože jeho dominantní hemisféra byla s největší pravděpodobností levá. Jedná se zkrátka o nepatřičný zásah do složitého procesu.
I přes existenci dominantní hemisféry jsou ale kontroly pohybů a záznamu senzorických informací schopny obě. Aby nedocházelo k interferenci a hemisféry pracovaly synchronně, existuje mohutný svazek komisurálních vláken – corpus callosum.
Při protětí corpus callosum dojde ke stavu, kdy postižený ztrácí kontrolu nad polovinou těla, respektive vzniknou dvě nezávislé vědomé hemisféry. Jsou dobře zdokumentovány případy, kdy si pacient holil pouze polovinu obličeje, česal si vlasy na každé straně zcela jinak nebo byl v extrémním případě napaden končetinou, kterou neovládal.
U osob s poškozenou nedominantní hemisférou bylo prokázáno, že dochází k poruše interpretace hudby, řeči těla a intonace jiných osob apod. Je tedy pravděpodobné, že nedominantní hemisféra je více využívána pro odlišné činnosti.
_
Řeč
Schopnost řeči je jednou z nejlépe prozkoumaných vyšších funkcí CNS. Existují dva aspekty řeči: senzorický aspekt, který využívá informace ze sluchové a vizuální kůry, a motorický aspekt, ten zapojuje motorické oblasti pro synchronizaci dýchacích svalů a mluvidel.
Senzorický aspekt
Jakmile komunikujeme s jinou osobou, jsou zvuky, které tvoří slova, zachyceny v primární sluchové kůře a v sekundární jsou rozpoznány zvukové motivy slov. Motivy jsou převedeny do Wernickeho oblasti, kde je vyhodnocen jejich význam. Wernickeho oblast také připraví myšlenku, kterou chceme sdělit, a vybere k tomu vhodná slova. Na tomto kroku se pravděpodobně menší měrou podílí i prefrontální kortex, neboť jeho poškození omezuje schopnost vyjadřování. Obsahově, lexikálně a stylisticky připravený obsah je pak přes fasciculus arcuatus převeden do Brocova centra.
Senzorický aspekt ve čteném textu probíhá totožně, pouze mezi vizuální kortex a Wernickeho centrum je vložen ještě g. angularis (viz výše).
Motorický aspekt
Brocovo centrum na základě informací z Wernickeho oblasti připraví motorický plán provedení a ten převede asociačními vlákny do motorické oblasti pro příslušné svaly. Tohoto kroku se účastní i mozeček, který určuje a hlídá přesné časování pohybů, a bazální ganglia.
Pomocí synchronních pohybů úst, jazyka, laryngu, hlasivek a dýchacích svalů pak přesně vokalizujeme myšlenku vzniklou ve Wernickeho centru.
_
Paměť
Vzpomínky jsou uchovány v mozku pomocí změn synaptického přenosu mezi neurony, ke kterým dochází v důsledku opakované a časté aktivace určité sekvence neuronů. Tyto změny mohou nabývat nejrůznějších podob, od modifikací již existujících spojů, např. mechanismem posílení nebo zeslabení odpovědi postsynaptického neuronu na stimulus nebo modifikacemi postsynaptické denzity, až ke vzniku zcela nových synapsí. Takto změněné synapse se nazývají paměťové stopy.
Paměťové stopy mohou vznikat v celém CNS, například v míše lze takto modulovat funkci reflexů. Dlouhodobé vzpomínky, které si spojujeme s intelektuálními funkcemi mozku, jsou ale uchovány převážně v kortexu.
Předpokládá se, že to, v jaké oblasti kůry nalezneme příslušnou paměťovou stopu, záleží především na jejím charakteru a vstupním senzorickém stimulu. Tak jsou informace o tvaru předmětů uloženy ve vizuálním kortexu a jejich názvy zase ve sluchové kůře.
Přestože si proces učení – tedy vznik nových paměťových stop – obvykle spojujeme s nabýváním informací, opak je pravdou. Většina senzorických impulsů je označena za nepotřebné a senzorické dráhy jsou modifikovány tak, aby daný stimulus ignorovaly. Tento osud potká více než 99 % příchozích senzorických informací. O tom, zda bude daná informace ignorována, rozhoduje z velké části limbický systém. Ten totiž senzorickým vstupům, které aktivují centra odměny nebo trestu, přiřadí tzv. afektivní kvalitu.
Pokud senzorická informace nemění aktivitu center odměny nebo trestu a není jí tak přiřazena afektivní kvalita, dochází k postupné ztrátě odpovědi na tento stimulus a korové oblasti jím přestávají být aktivovány. Tento jev se nazývá habituace a jedná se o typ negativní paměti. Pokud ovšem limbický systém přiřadí určitému senzorickému vstupu afektivní kvalitu a kůra se s podnětem setkává opakovaně, uvidíme naopak vzrůstající intenzitu odpovědi, jelikož dochází k facilitaci synaptických spojů. Říkáme, že odpověď byla posílena. Tento typ pozitivní paměti nazýváme senzitizace.
Kandel a jeho kolegové objevili u mořského plže aplazie mechanismus, který zajišťuje změny synaptických spojů. Zjistili, že na presynaptický terminál senzorického neuronu (někdy též označovaného jako senzorický terminál), přiléhá ještě jedno zakončení axonu. To se nazývá facilitační terminál a pochází ze serotoninergního neuronu. U aplazie je aktivován bolestivým stimulem. Předpokládá se, že u člověka mají neurony dávající vznik facilitačním terminálům úzkou souvislost s limbickým systémem. Tento terminál hraje významnou funkční úlohu v mechanismech habituace i facilitace.
Habituace
Habituace nastává, pokud je opakovaně aktivován presynaptický terminál senzorického neuronu bez současné aktivace facilitačního terminálu. Zprvu je postsynaptický excitační potenciál normální, ale s opakovanou stimulací se stále zmenšuje, až nakonec vyhasíná. Můžeme říci, že k aktivaci facilitačního terminálu dochází při přiřazení afektivní kvality. Jestliže stimulu není v limbickém systému tato kvalita přiřazena, nedojde k aktivaci facilitačního terminálu a senzorický terminál je habituován.
Na molekulární úrovni spočívá mechanismus habituace v progresivním uzavírání kalciových kanálů presynaptického terminálu. Příčinu uzavírání kanálů doposud neznáme. Jelikož je ale počet kvant uvolněného neurotransmiteru přímo úměrný koncentraci intracelulárního kalcia, dochází spolu s progresivním uzavíráním kanálů k výraznému snižování excitačního postsynaptického potenciálu.
Facilitace
Aktivuje-li senzorická informace limbický systém, je stimulován spolu se senzorickým terminálem i terminál facilitační. Ten uvolní na povrch senzorického terminálu serotonin, který přes své receptory aktivuje adenylátcyklázu. V této signalizační dráze jsou dále aktivovány cAMP dependentní proteinkinázy, jež fosforylují draslíkové kanály a tím je uzavírají. Ztráta permeability membrány pro draslík prodlužuje trvání akčního potenciálu, což nakonec vyústí v delší interval otevření vápníkových kanálů, vyšší koncentraci intracelulárního kalcia, vylití úměrně většího množství kvant neurotransmiteru a konečně posílení excitačního postsynaptického potenciálu.
Takto zvýšená odpověď může trvat až tři týdny, a to i bez další stimulace facilitačního terminálu.
Klasifikace paměti
Paměť můžeme klasifikovat na základě hlediska typu uchovávaných informací nebo doby, po kterou přetrvávají paměťové stopy.
Na základě typu informací rozlišujeme:
1) Deklarativní paměť (též explicitní)
2) Procedurální paměť (též implicitní)
Deklarativní paměť
Vyžaduje práci s paměťovou stopou na vědomé úrovni. Lze ji dále rozčlenit na sémantickou a epizodickou paměť.
Sémantická paměť obsahuje neosobní informace zbavené svého původního kontextu. Jedná se převážně o osvojené abstraktní vědomosti. Jako příklad sémantické paměťové stopy nám poslouží třeba věta: Praha je hlavní město České republiky. Typická abstraktní vědomost zbavená kontextu, který doprovázel její získání.
Epizodická paměť obsahuje informace s původním kontextem – vybavíme si místo a čas, kdy se daná událost stala, apod. Často jde o velmi osobní zážitky doprovázené emocemi. Velkou část epizodické paměti tvoří autobiografické vzpomínky.
Procedurální paměť
Nevyžaduje uvědomění paměťové stopy, rozumíme jí například nabyté motorické dovednosti. V procedurální paměti je tedy třeba uložena hra na kytaru nebo golfový švih. Procedurální paměťové stopy získáváme procesem motorického učení, které využívá bazálních ganglií a mozečku.
Na základě doby přetrvávání paměťových stop rozlišujeme:
1) Krátkodobá paměť (sekundy až minuty)
2) Střednědobá paměť (dny až týdny)
3) Dlouhodobá paměť (měsíce až roky)
Krátkodobá paměť
Krátkodobá paměť je typická svou schopností najednou uchovat maximálně 7 až 10 diskrétních informací – např. telefonní číslo. Pravděpodobně funguje na principu opakujících se neuronálních okruhů, ve kterých na vědomé úrovni udržujeme aktivitu. Kódovaná informace krouží v těchto okruzích a jejich aktivita vyhasne při ztrátě koncentrace.
Krátkodobou paměť nesmíme zaměňovat s operační pamětí, speciální funkcí prefrontálního kortexu. Liší se charakterem uchovávaných informací: diskrétní fakta (např. číslice) v krátkodobé paměti a složité abstraktní konstrukty, problémy nebo plány v operační paměti. Dalším rozdílem je, že problém uložený v operační paměti je z ní po svém vyřešení odstraněn.
Příklad rozdílu:
Operační paměť: Pokud myslíme na to, že je potřeba, abychom snědli snídani, umyli si zuby a odjeli do práce, bude plán „umýt si zuby” hned po svém splnění z operační paměti vymazán.
Krátkodobá paměť: Pokud si pamatujeme sekvenci 7-5-3-2-4-5, zůstane uložena tak dlouho, dokud se na ni budeme plně koncentrovat.
Střednědobá paměť
Střednědobé paměťové stopy přetrvávají až týdny a jsou ztraceny, nedojde-li k jejich opětovné aktivaci. Při častém opakovaném vybavování se z nich stávají dlouhodobé paměťové stopy. Má se za to, že ukládání vzpomínek do střednědobé paměti je dokonce zcela nutné pro vznik dlouhodobých paměťových stop. Mechanismem střednědobé paměti jsou habituace a facilitace popsané výše.
Dlouhodobá paměť
Neexistuje přesná hranice mezi udržovanou střednědobou paměťovou stopou a stopou dlouhodobou. Má se za to, že dlouhodobá paměť je uchovávána morfologickými změnami synapsí namísto změn fyzikálně-chemických, jako je tomu u habituace a facilitace.
Z experimentů vyplývá, že vzpomínka nemůže být uložena do dlouhodobé paměti, jestliže byly člověku podány látky blokující replikaci proteinů. Proto se předpokládá, že paměťová stopa je dlouhodobě uložena ve změnách morfologie presynaptického terminálu. Ty mohou být následující:
1) Zvětšení plochy aktivní zóny
2) Zvýšení počtu vezikul s neurotransmiterem, která jsou exocytována
3) Zvýšení počtu presynaptických terminálů daného neuronu
4) Vyvolání změn v postsynaptické denzitě, jež mění morfologii postsynaptického neuronu (např. vznik dendritických trnů).
Změny v síle synapse souhrnně označujeme pojmem synaptická plasticita.
Část těchto změn je vyvolána excesivním influxem kalcia do cytoplazmy neuronu, která aktivuje kalcium-kalmodulin dependentní kinázy a s nimi i příslušné signální dráhy.
Zajímavé je, jak změny v presynaptickém terminálu ovlivní terminál postsynaptický. Nejlépe prostudovaným vztahem v tomto typu synaptické plasticity je dlouhodobá potenciace (long-term potentiation – LTP) neuronů hipokampu. Její princip spočívá v dlouhodobém zesílení synaptického přenosu mezi dvěma neurony jako výsledek jejich současné stimulace.
Presynaptický neuron uvolňuje do synaptické štěrbiny glutamát, který se váže na AMPA receptory (AMPARs), nespecifické kationtové kanály s největší permeabilitou pro ionty sodíku. Kromě AMPARs se glutamát váže také na NMDA receptory (NMDARs). Jejich ústí je ale v klidovém stavu blokováno iontem magnesia. Až ve chvíli, kdy excitační postsynaptický potenciál z AMPARs dosáhne dostatečné hodnoty, je kation magnesia Coulombovskými silami z ústí NMDARs vytlačen. Teprve nyní se NMDARs projeví jako kationtový kanál a zvýší permeabilitu membrány pro vápenaté ionty (mimo jiné, jedná se o nespecifický kanál). Excesivní intracelulární kalcium aktivuje kalcium-kalmodulin dependentní kinázy a započne přestavba postsynaptické denzity. Nejdříve se pouze zvyšuje počet vystavených AMPARs a NMDARs. Později přibývá i strukturální složky postsynaptické denzity a na postsynaptickém terminálu začne vyčnívat drobný výstupek, který se s opakovanou aktivitou formuje v dendritický trn.
Konsolidace paměti
Konsolidace paměti je proces, který aktivuje chemické, fyzikální a morfologické změny synapsí tak, aby byla informace uložena do dlouhodobé paměti. Minimální čas pro konsolidaci paměťových stop činí 10 minut; čím déle vědomě nad vzpomínkou přemítáme, tím je konsolidace silnější.
Významné je, že nové paměťové stopy jsou kodifikovány, čili porovnány se starými vzpomínkami a na základě rozdílů a shody zařazeny do skupin k již existujícím vzpomínkám. To velmi napomáhá procesu rozpomínání.
Autor podkapitoly: Patrik Maďa
![]()