Náplň podkapitoly:
1. Úvod do somatosenzoriky
2. Mechanocepce
3. Vedení somatosenzorických informací
4. Bolest
_
Úvod do somatosenzoriky
Somatosenzorika je souhrnný název pro nervové mechanismy, které sbírají, vedou a vyhodnocují somatické vjemy z celého těla. Obecně lze říci, že somatické vjemy se člení do tří kategorií: mechanocepční, termocepční a nocicepční.
Mechanocepční vjemy
Mechanocepce je zprostředkována mechanoreceptory, jež v každém okamžiku zaznamenávají hmatové a polohové informace. Tyto receptory jsou stimulovány prostorovými změnami tkání, např. vzájemným pohybem sousedících vrstev tkání.
Termocepční vjemy
Termocepce je snímána termoceptory ve tkáních.
Nocicepční vjemy
Vnímání bolesti zprostředkovávají volná nervová zakončení, která jsou drážděna faktory poškozujícími tkáň.
V této kapitole se budeme blíže zabývat mechanocepcí a nocicepcí.
_
Mechanocepce
Základními mechanocepčními požitky jsou hmat, tlak a vibrace. Přestože často bývají považovány za odlišné smyslové vjemy, tak zcela to neplatí. Do jisté míry je registrují stejné receptory. Liší se pouze tím, že hmat je obvykle vnímán na povrchu kůže, kdežto tlak v hlubších tkáních (např. svalové fascie). Vibrační vjem pak vzniká opakovanou rychlou stimulací mechanoceptorů.
Mechanoceptory
Existují desítky různých mechanoceptorů často se lišících pouze nevýznamně. Tuto plejádu fyziologických senzorů lze rozdělit do šesti zcela odlišných typů:
1) Volná nervová zakončení
2) Meissnerova tělíska
3) Merkelovy disky
4) Receptory vlasového folikulu
5) Ruffiniho zakončení
6) Paciniho tělíska
Volná nervová zakončení
Volná nervová zakončení jsou difúzně rozmístěna v kůži i v dalších tkáních po celém těle. Obzvláště citlivá jsou na hmatový vjem, ale stejně jako ostatní receptory mají schopnost registrovat i tlak a vibrace. Obzvláště hustě jsou rozmístěna na oční rohovce jako jediný tamější typ mechanoceptoru.
Volná nervová vlákna také odpovídají za pocit svědění. Má se za to, že existují receptory, které jsou na vjem svědění funkčně specializovány a nacházejí se v povrchových vrstvách kůže. Morfologický rozdíl mezi nimi a obvyklými volnými zakončeními zjištěn nebyl. Evolučně svědění vzniklo jako obrana proti bodavému hmyzu živícímu se krví. Tento vjem informuje organismus o přítomnosti a lokalizaci takového hmyzu, a tak umožňuje zabránit napadení a následnému porušení kůže. U psů (a jiných psovitých šelem) existuje dokonce speciální mimovolní škrabací reflex spuštěný při podráždění volných nervových vláken v okrscích kůže, které jsou vzájemně vzdáleny o délku průměrného skoku blechy.
Meissnerova tělíska
Meissnerova tělíska jsou prodloužená obalená zakončení velkého myelinizovaného nervového vlákna. Uvnitř svého obalu se ještě dělí do drobných terminálních filament. Receptor je stimulován tak, že dojde k deformaci pouzdra a tím i ke stimulaci nervového zakončení.
Meissnerova tělíska jsou obzvláště hustě rozmístěná v neochlupených oblastech kůže, zejména pak na konečcích prstů. Obzvláštní citlivost vykazují na pohyb objektů po povrchu kůže a nízkofrekvenční vibrace.
Merkelovy disky
Tento typ receptoru se nachází po celé ploše kůže. V neochlupených oblastech doprovází Meissnerova tělíska, velmi se ale od nich liší způsobem signalizace, na rozdíl od nich jsou totiž Merkelovy disky méně citlivé na podráždění. Při podráždění sice zprvu vysílají silný signál, ten ale postupně slábne, až dosáhne určité udržovací. Díky této vlastnosti poskytují zejména informace o trvajícím kontaktu pokožky s objektem.
Pro lepší ilustraci rozdílné funkce Merkelových disků a Meissnerových tělísek si lze představit následující příklady:
1) Pokud slabě přejedeme prstem po nějaké tkanině, je tato informace snímána zejména (ne výhradně!) prostřednictvím Meissnerových tělísek.
2) Pokud v ruce držím tužku, je její kontakt s pokožkou neustále snímán Merkelovými disky.
Merkelovy disky se na některých místech pod epitelem kůže kupí do tzv. Iggo dómreceptorů. Tím se vytvářejí drobné hrbolky vystupující nad povrch kůže. Všechny Merkelovy disky tvořící jeden Iggo dóm jsou inervovány jediným velkým myelinizovaným vláknem. Iggo dómy mají oproti jednotlivým diskům vysokou citlivost. Slouží proto zejména k analýze povrchu objektu, který se dostal do kontaktu s kůží.
Receptory vlasového folikulu
Sestávají z nervového vlákna, jež obtáčí vlasový folikul. Fungují jako ekvivalent Meissnerových tělísek v ochlupených lokalitách kůže. Registrují zejména pohyb objektu po povrchu těla.
Ruffiniho tělíska
Jde o rozvětvená obalená nervová zakončení. Snímají zejména deformaci tkání, proto jsou obzvláště citlivá na tlak. Nacházejí se i ve tkáních přiléhajících ke kloubům. Díky své schopnosti registrovat deformace mohou snímat např. úroveň rotace v daném kloubu apod.
Paciniho tělíska
Nacházejí se jak na povrchu těla, tak v hlubokých tkáních. Vyznačují se schopností téměř okamžité adaptace (v setinách sekund), proto je může stimulovat pouze velmi rychle se měnící mechanický podnět (např. komprese). Tím jsou obzvláště cenné v registraci vibrací o vyšší frekvenci.
Detekce vibrací závisí tedy na jejich frekvenci. Vysokofrekvenční vibrace snímají Paciniho tělíska, nízkofrekvenční Meissnerova tělíska. Detekce se ovšem vždy více či méně účastní všechny typy mechanoreceptorů.
_
Vedení somatosenzorických informací
Všechna vlákna, která vedou senzorické informace vstupují do zadního rohu míšního. Dále jsou vedena jednou ze dvou alternativních cest, a to buď systémem zadních provazců amediálního lemnisku, nebo anterolaterálním systémem. Tyto dva systémy se pak opět setkávají až na úrovni thalamu.
Systém zadních provazců a mediálního lemnisku
Do tohoto systému vstupují zejména velká myelinizovaná nervová vlákna, která pocházejí z pseudounipolárních neuronů, jejichž těla jsou uložena ve spinálních gangliích (1. neuron dráhy). Ascendentní senzorická vlákna stoupají ze zadního rohu míšního v zadních míšních provazcích. Kříží se na přechodu míchy a kmene. První neurony končí na neuronech nc. gracilis a nc. cuneatus.
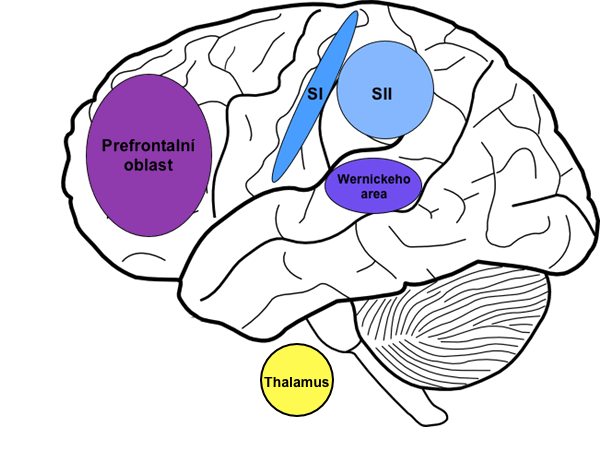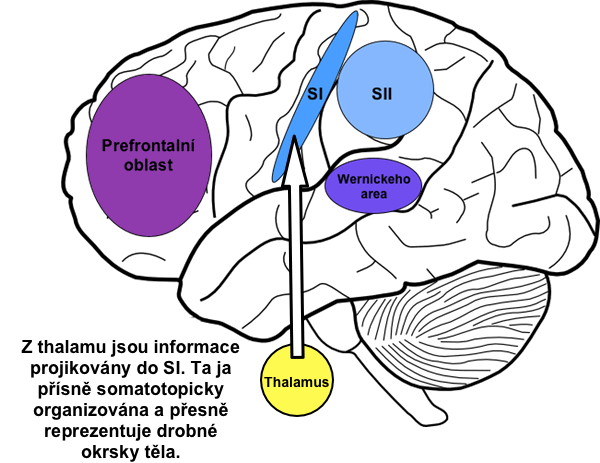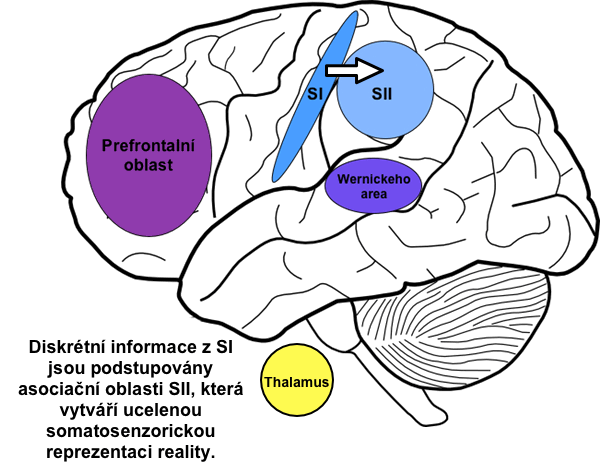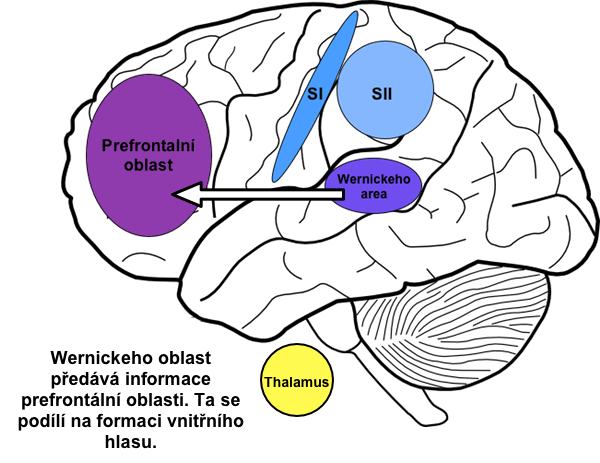
Prostřednictvím tohoto systému jsou vedeny mechanocepční vjemy. Je pro něj typická velmi přísná somatotopická organizace, což znamená, že neuron na kterékoli úrovni tohoto systému je přiřazen dané oblasti těla (např. části dermatomu). Stav receptorů v této oblasti svou aktivitou reprezentuje danou oblast těla, proto můžeme přesně lokalizovat daný podnět. Například víme, kterým prstem se dotýkáme objektu.
Anterolaterální systém
V tomto systému je informace vedena zejména menšími nervovými vlákny bez myelinu. Ihned po vstupu do příslušného míšního segmentu se první senzorický neuron přepojí v dorzálním rohu míšním. Poté vstoupí do kontralaterálního předního nebo laterálního míšního svazku. To mimo jiné znamená, že informace vedené anterolaterálním systémem se kříží již na úrovni příslušného segmentu. Druhý neuron končí na neuronech v mozkovém kmeni a thalamu.
Tímto systémem jsou vedeny vjemy bolesti, termické počitky a svědění. Somatotopická organizace zde není přísně dodržena, lokalizace podnětu je tedy obtížnější.
Somatosenzorické oblasti kůry
Z thalamu je informace vedena do kůry cestou tr. thalamocorticalis. Axony thalamických neuronů končí za sulcus centralis na g. postcentralis. Tato oblast kůry se nazývá somatosenzorická area I (SI), někdy též somatosenzorický kortex.
SI je přísně somatotopicky organizována a její jednotlivé úseky můžeme přiřadit k částem těla. O této místní reprezentaci často hovoříme jako o somatosenzorickém homunkulovi, neboť můžeme na povrch g. postcentralis promítnout zmenšenou lidskou postavu tak, aby daná část těla ležela na své místní reprezentaci. Tato postavička však je značně deformovaná, neboť okrsky kůry jsou proporční vzhledem k úrovni inervace, nikoli k reálné velikosti části (homunkulus má například největší ruce).
Při poškození této oblasti dochází k několika jevům:
1) Osoba není schopna lokalizovat vjem. Např. se zavřenýma očima nedokáže říci, které dolní končetiny se lékař dotýká.
2) Není schopna rozlišit lehký dotek od silného tlaku.
3) Není schopna posoudit váhu předmětů, které drží.
4) Není schopna posoudit tvary předmětů (tento nález se nazývá astereognozie).
5) Nerozlišuje texturu objektů.
Existuje i somatosenzorická area II (SII), která naléhá na dorzobazální část SI. Jelikož je velmi špatně prozkoumaná, o její funkci toho mnoho nevíme. Většina vstupů do SII nepřichází z thalamu, ale z SI a dalších oblastí kůry (vizuální, sluchové, apod.). Mohlo by se tedy jednat o asociační oblast, která se podílí na integraci jednotlivých senzorických vjemů a spolutvoří tak vnitřní reprezentaci reality. Každopádně poškozením této oblasti nevzniká žádná závažná patologie.
Na zpracování senzorických informací se podílejí i další dvě asociační oblasti: area 5 a area 7. Ty získávají vstupy z SI a SII a interpretují senzorické informace. Stimulací těchto oblastí lze vyvolat vjemy, jež daná osoba může interpretovat tak, že např. v levé ruce drží gumový míček apod.
_
Bolest
Bolest je fyziologický vjem sloužící jako ochranný mechanismus, její úkol spočívá v zabránění dalšímu poškození tkáně. Pokud například člověk leží dlouhodobě bez hnutí, vlivem tlaku podložky na tělo dochází ke kompresím, které dávají vznik lokální ischemii kůže. Ta pak může vést až k proleženinám – dekubitům. Fyziologicky však u zdravého jedince k tomuto jevu nedojde, neboť ischemie tkáně u něj vyvolá vjem bolesti a on změní polohu. Proto dekubity pozorujeme převážně u imobilních pacientů, kteří nedokážou volně měnit polohu, nebo u osob, jež ztratily schopnost cítit bolest.
Typy bolesti
Existují dva základní typy bolesti: pomalá a rychlá. Liší se rychlosti vzniku a rozvoje.
Rychlá bolest
Rychlá bolest se vyvíjí během desetin sekundy. Bývá také označována jako ostrá bolest. Tento vjem nejčastěji vzniká při mechanickém porušení kožního krytu typu bodnutí nebo říznutí, u akutně vzniklých popálenin, při zásahu elektrickým proudem apod. Naopak nevzniká v hlubokých tkáních těla.
Pomalá bolest
Pomalá bolest nastupuje až po několika sekundách od působení bolestivého stimulu. Pomalu se rozvíjí a zvyšuje svou intenzitu. Je vždy spojená s destrukcí tkáně, chronickými stavy a může vést k významnému snížení kvality života. Nastává jak v hlubokých, tak v povrchových tkáních.
Nociceptory
Recepory bolesti neboli nociceptory jsou volná nervová zakončení rozvrstvená po celém těle – v kůži, v periostu, ve stěnách velkých arterií, v kloubech atd. Nociceptory reagují na podněty, jež lze rozdělit do tří typů: mechanické, chemické a termické. Všechny tři typy dovedou vyvolat pomalou bolest, ale pouze mechanické a termické vyvolají bolest rychlou.
Chemické podněty, které stimulují nociceptory, jsou jednak látky úzce spjaté se zánětem (bradykininy, serotonin, histamin, proteolytické enzymy a kyselé prostředí – respektive vodíkové ionty) a jednak látky uvolněné při rozpadu buněk (draselné ionty, acetylcholin apod.).
Je zapotřebí připomenout, že draselné ionty vyvolají pomalý typ bolesti pouze ve vysoké koncentraci, které bez rozpadu buněk nelze jinými endogenními mechanismy dosáhnout. Po dosažení této „kritické úrovně” je intenzita bolesti úměrná koncentraci. S dalším rozpadem buněk a uvolňováním draslíku se tedy zvyšuje intenzita bolestivého vjemu.
Nociceptory mají ještě jednu velmi důležitou vlastnost. Na rozdíl od jakýchkoliv jiných receptorů se nedokážou adaptovat. Říkáme proto, že receptory bolesti jsou neadaptivní. Tato vlastnost zaručuje, že bolestivému vjemu bude věnována pozornost a bude neustále vědomě zpracováván. Na druhou stranu také odpovídá za nízkou kvalitu života pacientů s chronickou bolestí. Citlivost nociceptorů se dokonce za určitých podmínek zvyšuje a vjem bolesti je čím dál tím intenzivnější. Toto zvyšování citlivosti nazýváme hyperalgesie.
Vedení bolestivého vjemu
Při stimulaci nociceptoru je bolest vedena periferními nervovými vlákny v závislosti na typu bolesti. Rychlou bolest přenášejí silné myelinizováné axony, pomalou bolest naopak tenká nemyelinizovaná vlákna, která díky těmto vlastnostem přenášejí vzruch i 20x pomaleji.
Kvůli tomuto dvojímu systému vedení často dochází ke zdvojení bolestivého vjemu. Jedná se o jev fyziologicky poměrně významný, neboť rychlá bolest upozorní na okamžité poškození tkáně doprovozené jen slabou pomalou bolestí. Pokud však poškozující stimulus přetrvává, zvyšuje se i intenzita pomalé bolesti a nutí daného jedince dále vyhledávat způsob úlevy.
Jakmile nervová vlákna dosáhnou příslušného míšního segmentu, pokračují do vyšších etáží CNS jednou ze dvou drah: tr. neospinothalamicus nebo tr. paleospinothalamicus.
Neospinothalamický trakt
Neospinothalamický trakt vede převážně vjemy rychlé bolesti. Neurony pro vedení rychlé bolesti (první neurony) končí v Rexedově zóně I (lamina marginalis) dorzálních rohů míšních. Tam excitují druhé neurony neospinothalamického traktu, jež se okamžitě v daném segmentu kříží před canalis centralis a vstupují do předních a laterálních svazků míšních, kterými pokračují vzhůru do vyšších etáží jako část anterolaterálního systému.
Malá část vláken končí již v retikulární formaci kmene. Bolestivý stimulus tak zvyšuje bdělost prostřednictvím zvýšení aktivity v ascendentním retikulárním aktivačním systému (ARAS). Tím udržuje pozornost vůči bolestivému vjemu.
Většina vláken však končí až na ventrobazálním komplexu jader thalamu. Zde končí neospinothalamický trakt a jeho druhé neurony jsou přepojeny na tr. thalamocorticalis, který vede bolest do SI, čímž umožní její lokalizaci. Kromě výše uvedeného se přepojuje i na další dráhy, jež rozvedou bolest do bazálních oblastí mozku a limbického systému.
Paleospinothalamický trakt
Paleospinothalamický trakt vede vjemy pomalé bolesti. Drobné neurony, jejichž nemyelinizovaná vlákna přenášela pomalý typ bolesti, končí v Rexedových zónach II a III (souhrnně substantia gelatinosa) zadních rohů míšních. Pak je bolestivý vjem několikrát přepojen na interneuronech zadního rohu (tím dochází k významnému zpoždění vjemu), až dosáhne Rexedovy zóny IV. Poté se kříží před canalis centralis a připojuje se k vláknům pro rychlou bolest v anterolaterálním systému.
Většina vláken končí již v mozkovém kmeni a to na:
1) Retikulární formaci kmene
2) Area tectalis
3) Periakveduktální šedi (obklopuje aquaductus Sylvii)
Pouze desetina vláken pokračuje výše do thalamu. To spolu s násobným přepojením už v zadním rohu míšním vysvětluje, proč se pomalá bolest špatně lokalizuje. Jednak do vyšších etáží nepřichází dostatek informací pro přesnou lokalizaci, jednak první neurony končí na interneuronech, které signál přepojují i na vlákna vedoucí do jiných míšních segmentů.
Není bez zajímavosti, že bolest je jediný senzorický vjem, na jehož uvědomění není třeba aktivita kůry. Dosažení úrovně thalamu je dostatečné pro vědomé zpracování bolesti.
Analgetické systémy CNS
CNS má omezenou schopnost potlačit bolestivé vjemy. Děje se tak prostřednictvím aktivace tzn. analgetického systému, který se skládá ze tří hlavních částí:
1) Periakveduktální šedi
2) Nc. raphe magnus
3) Bolest inhibujícího komplexu zadních rohů míšních
Pokud je zachycen silný bolestivý vjem, dojde automaticky i ke stimulaci periakveduktální šedi. Její neurony vysílají vlákna do nc. raphe, kde uvolňují enkefalin (jeho přesná úloha v nc. raphe není popsána). Descendentní projekce ze serotoninergních neuronů nc. raphe končí na komplexu interneuronů v zadních rozích míšních. Tyto inhibiční interneuronybrání přepojení vzruchu na neurony vedoucí bolestivý podnět anterolaterálním systémem. Děje se tak zejména prostřednictvím enkefalinu, jejž neurony v zadních rozích míšních sekretují. Má se za to, že enkefalin odpovídá jednak za presynaptickou, jednak za postsynaptickou inhibici prvních neuronů jak paleospinothalamického, tak i neospinothalamického traktu.
Autor podkapitoly: Patrik Maďa
![]()