Náplň podkapitoly:
1. Úvod do neurotransmisních systémů
2. Glutamánergní systém
3. GABAergní systém
4. Cholinergní systém
5. Katecholaminy jako neurotransmitery
6. Serotoninergní systém
7. Glycinový systém
8. Histaminergní systém
9. Purinergní neurotransmitery
10. Peptidy
11. Endogenní kanabonoidy
_
Úvod do neurotransmisních systémů
Neurotransmiter je látka, která je uvolněna neuronem ke specifické cílové buňce (respektive buňkám) a vyvolá v ní (nich) specifickou odezvu. Tímto cílem mohou být jiné neurony nebo orgány, zejména žlázy a svaly.
Na rozdíl od endokrinní signalizace je neurotransmise cílená na buňky v blízkém okolí zakončení neuronu, který látku uvolňuje, ať už do synaptické štěrbiny nebo do okolí tzv. varikozit (buttons en passant) v autonomním nervovém systému.
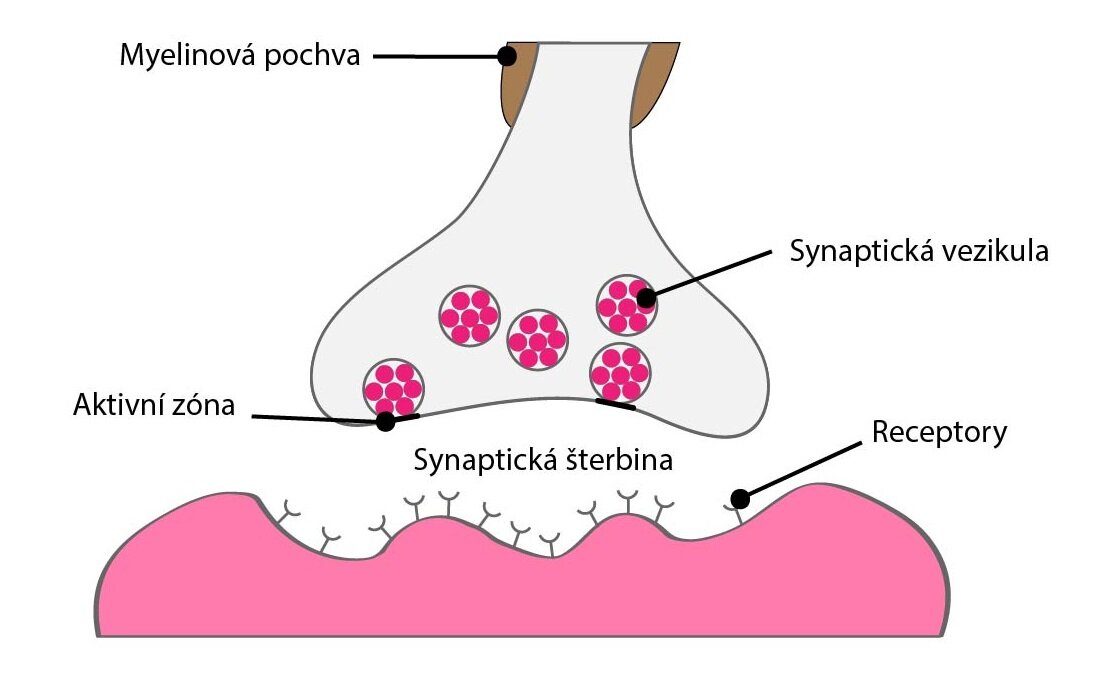
Rozdílem oproti parakrinní signalizaci je to, že neurotransmiter účinkuje nejen na cílové buňky, ale i na samotný neuron, který jej uvolnil. Tento fenomén většinou moduluje další vyplavování neurotransmiteru nebo jeho samotnou syntézu.
Další významné specifikum představuje skutečnost, že vazba neurotransmiteru a receptoru je přechodná, trvá od několika milisekund až po několik minut. Na druhou stranu změny, jež vyvolá, mohou být stejně pomíjivé, nebo naopak trvat i dny až týdny.
Okamžité účinky jsou obvykle zprostředkovány iontropními receptory, které samy zastávají funkci iontových kanálů (nebo jsou s nimi spřaženy) a vedou ke změně membránového potenciálu, případně k překročení depolarizačního prahu a vyvolání akčního potenciálu.
Dlouhodobé změny cílových buněk jsou zprostředkovány vazbou neurotransmiteru na metabotropní receptor, ten obvykle přes G-protein aktivuje intracelulární signalizační kaskády, jež vyvolají změnu exprese různých genů nebo aktivaci již preexistujících enzymů (např. cyklus fosforylace/defosforylace).
Koncept neurotransmise jako intercelulární komunikace neuronů s jiným neuronem nebo efektorovým orgánem již dnes není zcela platný. Ukazuje se, že některé gliové buňky, zejména astrocyty, jsou schopny neurotransmiter syntetizovat, a hlavně ho dokážou samy uvolňovat ke svým cílovým buňkám, stejně tak jako exprimovat na své membráně příslušné receptory.
Existuje obrovská plejáda látek, které se nacházejí v nervovém systému a které jsou syntetizovány neurony. Jen některé však mohou být považovány za neurotransmitery, a to pouze takové, jež splňují nasledující čtyři kritéria:
1) Látka je syntetizována presynaptickým neuronem
2) Látka je skladována v presynaptickém zakončení a uvolněna v dostatečném množství pro vyvolání změny v postsynaptickém neuronu nebo efektorovém orgánu
3) Pokud je tato látka podána exogenně, vyvolá stejnou reakci jako její endogenní uvolnění
4) Existuje specifický mechanismus pro inhibici účinku dané látky v synaptické štěrbině
Z chemického hlediska nejsou neurotransmitery jednotnou skupinou látek, jejich struktura je velmi různorodá. Obecně je můžeme rozdělit do dvou velkých skupin:
1) Velké molekuly
a) Peptidy: beta-endorfin, leu-enkefalin či substance P
b) Endogenní kanabinoidy
2) Malé molekuly
a) Aminokyseliny: glutamát, aspartát či glycin
b) Deriváty aminokyselin: GABA, katecholaminy (noradrenalin a dopamin) a serotonin
c) Acetylcholin
d) Ostatní: puriny (ATP, ADP, adenosin) nebo plyny (NO)
Obecně se dá říci, že skrze hematoencefalickou bariéru do CNS prostupují pouze prekurzory neurotransmiterů, nikoli hotové neurotransmitery. Toto poznání mělo velký význam pro léčbu některých neurologických onemocnění.
Syntéza většiny neurotransmiterů probíhá přímo v synaptickém zakončení. Výjimku představují peptidy, které se tvoří v těle, trofickém segmentu, neuronu.
_
Glutamatinergní systém
Glutamát je hlavní excitační neurotransmiter v CNS. Je v podstatě všudypřítomný, a nedá se proto hovořit o diskrétních centrech a jejich projekcích.
Má nezastupitelnou úlohu v tzv. synaptické plasticitě, tedy zániku starých, nevyužitých synapsí, posílení jiných synapsí nebo vzniku úplně nových. Tyto změny, které z velké části zprostředkovává glutamatinergní systém (ne ale výhradně), mohou v podstatě měnit interneuronální propojení v CNS z minuty na minutu.
Má se za to, že právě synaptická plasticita je neurobiolgický jev stojící za uchováváním vzpomínek a procesem učení.
Syntéza a inaktivace glutamátu – cyklus neuron-astrocyt
Jak již bylo uvedeno v úvodu kapitoly, hematoencefalická bariéra je nastavena tak, že glutamát z krve prakticky nikdy neprostupuje do CNS. Za normálních okolností probíhá téměř jen přesun glutamátu z CNS do krve.
Většina glutamátu je v neuronech produkována z glutaminu (enzym glutamináza), který sám o sobě vzniká v astrocytu. Z nich je transportován do neuronů. Pro tvorbu glutamátu (a současně i GABA) je zcela klíčová úzká kooperace neuronů a k nim přilehlých astrocytů.
Po uvolnění glutamátu do synaptické štěrbiny je jen jeho malá část vychytána zpět do presynaptického neuronu. Většina je naopak vychytána astrocytem, přenos zprostředkovávají glutamátové transportéry. U lidí se jedná o tzv. transportéry excitačních aminokyselin EAAT1 a EAAT2 (angl. Excitatory Amino Acid Transporter).
V astrocytu je vychytaný glutamát přeměněn na glutamin. Tato konverze je katalyzována glutaminsyntetázou a vyžaduje ATP a amoniak. Glutamin je zpět předán do neuronu, kde dochází k recyklaci glutamátu, který může být opět vyloučen do synaptické štěrbiny.
Klinická korelace:
U některých onemocnění roste koncentrace amoniaku v krvi – např. u onemocnění jater. Amoniak jako neutrální molekula snadno prostupuje hematoencefalickou bariérou. V přítomnosti glutamátu, kterého je v CNS mnoho, dochází k navázání amoniaku na glutamát za vzniku glutaminu. Současně se ale spotřebovalo ATP. Pokud je amoniaku velký nadbytek, vzniká deplece ATP v CNS a současně dochází k dysregulaci systémů dvou nejpočetnějších neurotransmiterů – glutamátu a GABA. Hovoříme o tzv. jaterní encefalopatii. Zvýšená koncentrace amoniaku v krvi je jen jedním z mechanismů, jak onemocnění jater ovlivňuje funkci CNS.
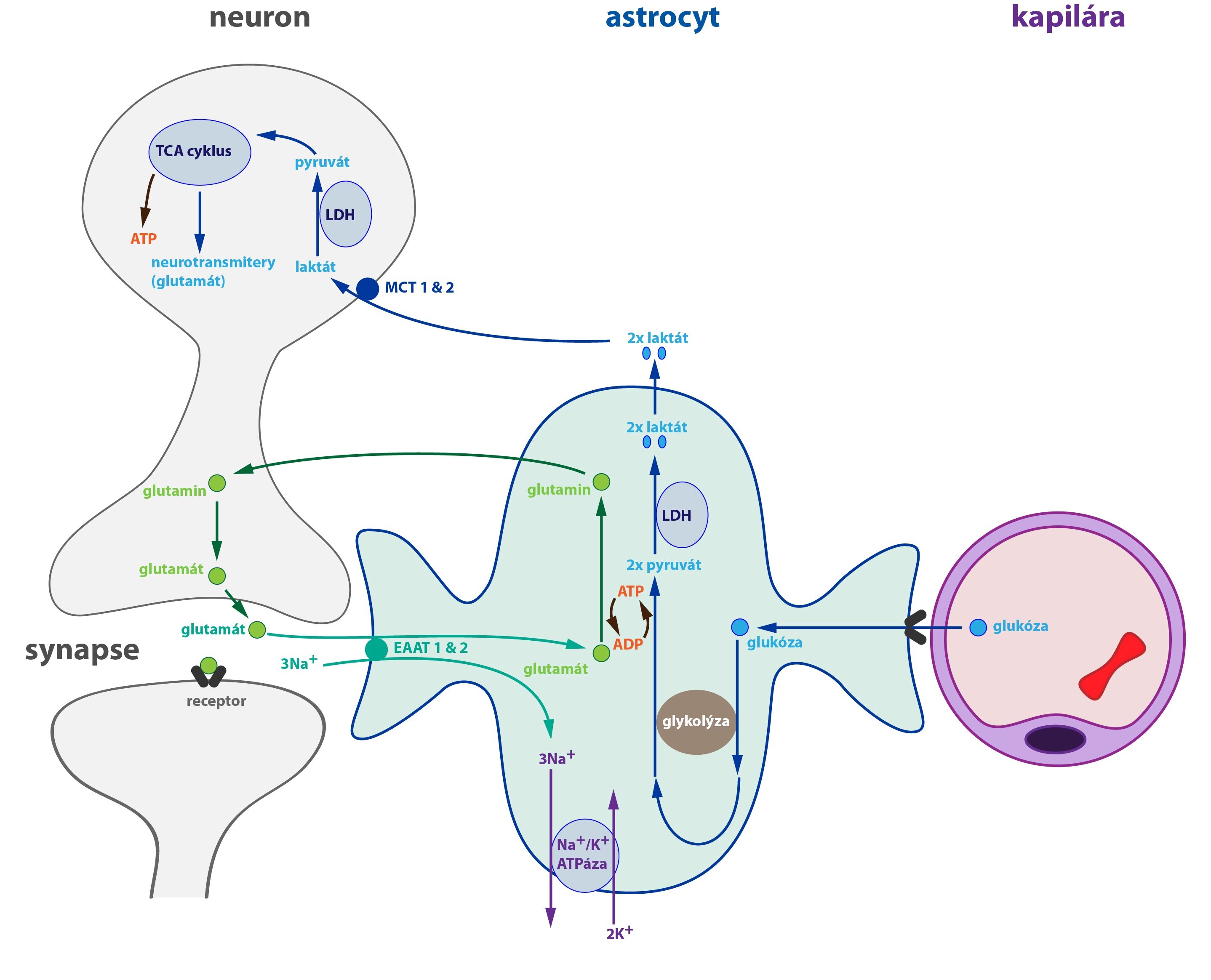
Nabízí se zajímavá otázka, proč astrocyt recykluje glutamát takto komplikovaně a proč nepředává neuronu přímo glutamát. Jedním z potenciálních vysvětlení je, že glutamát je neurotransmiter, a kdyby astrocyt jen tak volně vypouštěl glutamát do extracelulární tekutiny, vyvolával by odpověď – excitaci neuronů. Aby k tomuto nechtěnému ději nedocházelo, je glutamát nejdříve přeměněn na glutamin a až ten je předán neuronu. Poslední poznatky nicméně ukazují, že astrocyt za určitých okolností glutamát skutečně vypouští. Činí to ale vysoce cíleně.
Existuje několik receptorů pro glutamát, které se nazývají dle svého nejvýznamnějšího agonisty:
1) AMPA – receptory
2) NMDA – receptory
3) Kainátergní receptory
4) Metabotropní receptory (mGluR1-8)
1) AMPA – receptory (AMPARs)
Jedná se o ionotropní receptor aktivovaný ligandem. Při jeho otevření dochází k influxu vápníku, sodíku a exfluxu draslíku. Po aktivaci se velmi rychle zavírá.
2) NMDA – receptory (NMDARs)
Je také ionotropním receptorem, který zvyšuje permeabilitu membrány pro vápník, sodík a draslík. Narozdíl od AMPA – receptoru je uzavřen nejen proteinovou doménou, která změnou své konformace otevírá ústí kanálu, ale i hořečnatým kationtem. Kanál se uvolní pouze tehdy, dojde-li k dostatečné depolarizaci membrány (tu obvykle zprostředkují AMPARs) a vypuzení kladně nabitého iontu z kanálu prostými Coulombovými silami.
NMDARs stojí za influxem dostatečného množství kalcia na aktivaci kalcium-dependentních enzymů, které pak jsou schopny změnit vlastnosti postsynaptické denzity, hustotu receptorů na membráně apod. Jedná se tedy o jedny z hlavních faktorů podílejících se na synaptické plasticitě.
3) Kainátergní receptory
Kainátergní receptory (neboli kainátové receptory) jsou také inotropní receptory funkčně shodné s AMPARs.
4) Metabotropní receptory
Existuje jich 8 typů a značí se mGluR1-8. Nalézají se na presynaptických neuronech, jejichž aktivitu tlumí, nebo zvyšují obrat fosfatidylinositolu.
_
GABAergní systém
Kyselina gamma-aminomáselná je hlavním inhibičním neurotransmiterem v CNS. Je ve velkých koncentracích přítomná ve většině inhibičních neuronů mozku obratlovců, a není tak možné hovořit o diskrétním systému, kde větší nakupení GABAergních neuronů mají své aferenty a eferenty.
Syntéza a inaktivace GABA – cyklus neuron-astrocyt
Většina GABA vzniká dekarboxylací glutamátu (enzym glutamátdekarboxyláza). Stejně jako v případě glutamátu je i zde hlavním prekurzorem glutamin, který se do neuronu transportuje z astrocytu. Glutamin se v neuronu přeměňuje na glutamát (enzym glutamináza), ze kterého dekarboxylací vzniká GABA.
Ze synaptické štěrbiny je GABA vychytávána převážně astrocytem, kde se GABA přeměňuje na glutamin. Prvním krokem této přeměny je transaminace, kdy je aminoskupina nahrazena aldehydovou skupinou – vzniká sukcinátsemialdehyd (enzym GABAtransamináza). Následuje oxidace sukcinát semialdehydu na sukcinát, jenž se zapojí do KC. Ze sukcinátu může v KC vzniknout alfa-KG, který je dále přeměněn na glutamát (transaminace, aminace). Za spotřeby ATP a amoniaku je glutamát převeden na glutamin, který je přenesen do neuronu. Cyklus se uzavírá.
Existují 3 typy GABAergních receptorů:
1) GABAA
2) GABAB
3) GABAC
1) GABAA
Jedná se o ionotropní receptor, který zvyšuje permeabilitu pro Cl– ionty. Jeho aktivace vede k influxu Cl– iontů do cytoplazmy, kde díky svému zápornému náboji snižují membránový potenciál.
Je součástí tzv. supramolekulárního receptorového komplexu, který se skládá kromě GABAA také z receptorů pro benzodiazepiny, barbituráty, kortikosteroidy a alkohol. Tyto jednotlivé receptory tvoří ve svém středu kanál a všechny se podílejí na jeho aktivitě. Všechna vazebná místa tohoto supramolekulárního receptorového komplexu působí inhibičně na postsynaptický neuron, a to buď přímo (otevření kanálu) nebo nepřímo (potenciace působení GABA na GABAA).
2) GABAB
Jde o metabotropní receptor spřažený s Gi-proteinem, který tlumí aktivitu adenylátcyklázy. Snížení množství cAMP způsobí změny membránových proteinů (ve smyslu defosforylace/fosforylace) vedoucí k výraznému zvýšení propustnosti pro K+ (hyperpolarizace membrány), a naopak snížení aktivity kanálů pro Ca2+. Suma těchto změn v konečném důsledku snižuje množství neurotransmiteru vyplaveného cílovým neuronem.
Na tento typ receptorů jsou bohaté různé oblasti kortexu, thalamus a mozeček.
3) GABAC (nověji GABAA-rho receptor)
Jsou to ionotropní receptory spřažené s Cl– kanálem. Narozdíl od GABAA receptorů se pomalu otevírají, ale otevřené zůstávají mnohem déle. Jejich inhibiční účinek je tak asi 10x mocnější.
_
Cholinergní systém
Acetylcholin je jediný neurotransmiter, který má chemickou strukturu kvarterní amoniové sloučeniny, tedy odvozenou od aminu, jeho prekurzorem není žádná aminokyselina.
Je neurotransmiterem všech nervosvalových plotének obratlovců, všech presynaptických neuronů v autonomním nervovém systému, stejně jako všech postgangliových neuronů parasympatiku. V CNS moduluje celou řadu kortikálních aktivit, zejména bdění, spánek a konsolidaci paměťových stop.
Syntéza a inaktivace acetylcholinu
Acetylcholin je ester cholinu a acetátu. Syntetizuje se v jednom kroku – přenosem acetylu z AcCoA na cholin, reakci katalyzuje cholinacetyltransferáza.
Mozek získává téměř veškerý cholin z krve. Hlavním místem tvorby cholinu jsou játra, kde vzniká trojnásobnou methylací etanolaminu (donorem methylových skupin je SAM). Ethanolamin vzniká dekarboxylací serinu.
Ukončení cholinergní transmise zajišťuje enzym ze skupiny serinových hydroláz – acetylcholinesteráza (AChE), která je vázána na postsynaptické membráně. Produkty reakce jsou cholin (zpětně transportován do presynaptického neuronu, kde může být využit k další syntéze) a acetát. Pro katalytickou funkci AChE je klíčová triáda aminokyselin – serin, histidin a glutamát (podobná triáda se vyskytuje i u serinových proteáz, kde se ale namísto glutamátu vyskytuje aspartát) – hydroxylová skupina serinu napadá esterovou vazbu. AChE není substrátově zcela specifická, hydrolyzuje i jiné estery cholinu.
Působení acetylcholinu v nervovém systému je zprostředkováno dvěma typy receptorů:
1) Muskarinové receptory
2) Nikotinové receptory
1) Muskarinové receptory
Jde o metabotropní receptory spřažené s G-proteinem, který řídí iontové kanály. Odpověď postsynaptického terminálu na stimulus je z toho důvodu relativně pomalá. Zatím bylo objeveno pět typů muskarinových receptorů M1-5.
a) M1 receptory
M1 receptory, tzv. „neuronální”, se nacházejí ve velké denzitě v CNS, zejména pak v hipokampalní formaci a kůře. Jedná se o receptor zprostředkující excitační reakci prostřednictvím Gq-proteinu napojeného na signalizační kaskádu, jejímž konečným výsledkem je snížení permeability membrány pro K+. Má se za to, že oblenění jejich funkce nebo snížení jejich denzity je jednou z příčin demence.
b) M2 receptory
M2 receptory, tzv. „kardiální”, jsou nejvíce exprimovány na kardiomyocytech, ale ve velkých denzitách je najdeme i v neuronálních tkáních. Zprostředkují inhibiční působení prostřednictvím Gi-proteinu, který snížením intracelulární koncentrace cAMP aktivuje kanály pro K+ (exflux kalia vede k hyperpolarizaci). Tímto mechanismem působí n. vagus negativně chronotropně na sinuatriální uzel a negativně dromotropně na atrioventrikulární uzel. V CNS se vyskytují na presynaptických terminálech a jako autoreceptory zpětnovazebně tlumí vyplavování acetylcholinu v kůře a hipokampální formaci.
c) M3 receptory
M3 receptory, tzv. „žláz a hladkých svalů”, zprostředkují cholinergní stimulaci sekrece žláz a kontrakci hladkého svalstva GIT (a jiných orgánů). Tyto receptory jsou spřaženy s Gq-proteinem a prostřednictvím fosfolipázy C (katalyzuje vznik IP3 a DAG) zvýší intracelulární koncentraci kalcia. Přestože se v CNS vyskytují jen v malé denzitě, jsou schopny indukovat velmi silný emetický efekt.
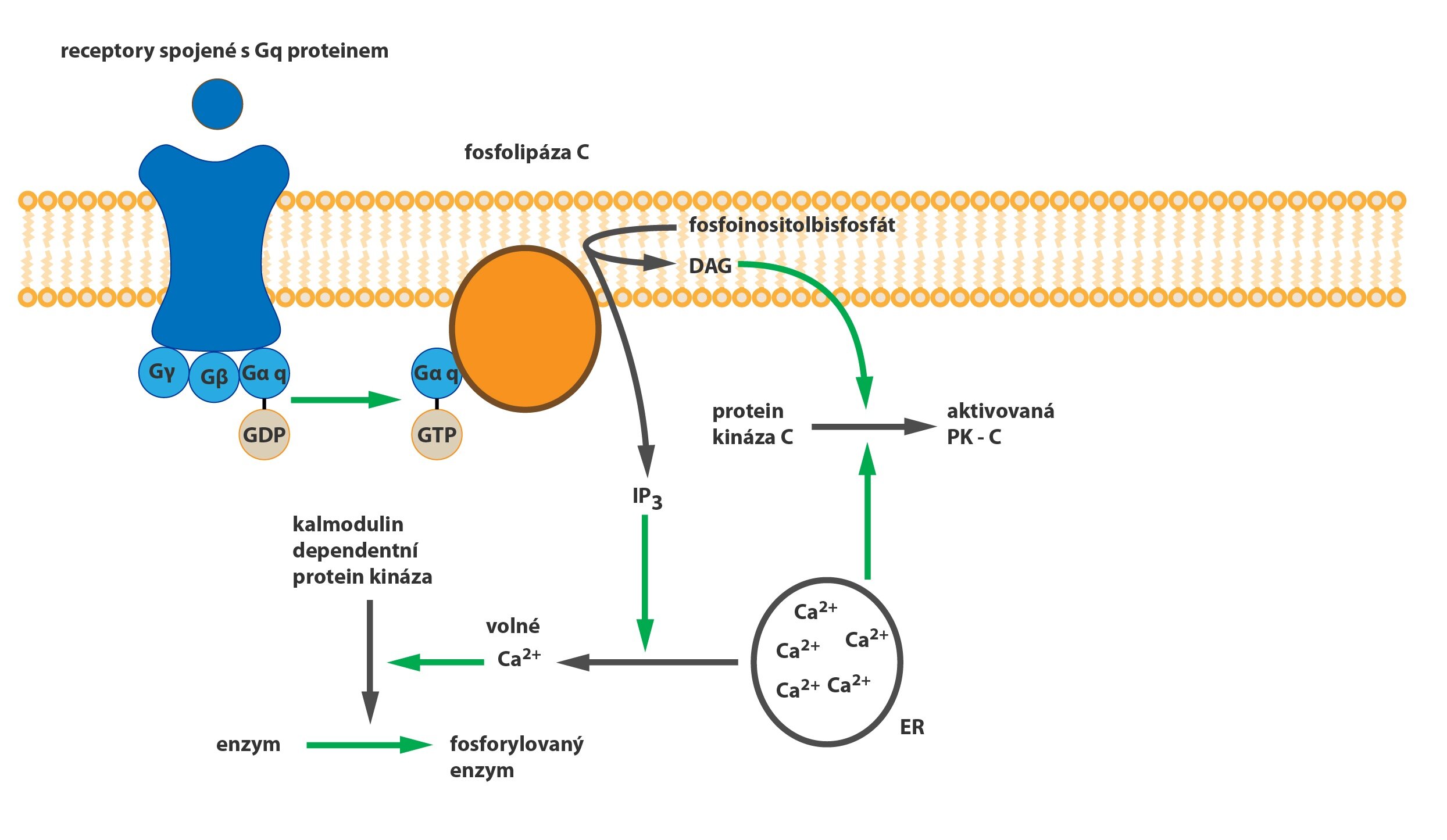
Zajímavý je účinek acetylcholinu na stěnu cév. Přestože acetylcholin působí kontrakci hladkého svalu, u cév pozorujeme vazodilatační účinek. Nejde o přímý účinek na svalovinu, ale spíše o heterotropní inhibici noradrenergní sympatické aktivity a stimulaci endotelu k produkci NO.
Receptory M3 jsou velmi intenzivně zkoumány kvůli své roli v patogenezi diabetu 2. typu. V CNS se nacházejí na strukturách, které odpovídají za monitoring a udržení hladiny glykemie (a tím i regulaci sekrece inzulinu), tedy v některých částech hypotalamu a parasympatických jádrech kmene. V pankreatu jsou ve velkých denzitách exprimovány na beta buňkách Langerhansových ostrůvků. Toto je významné, neboť některá antipsychotika používaná pro léčbu schizofrenie blokují M3 receptory a vedou tak dysregulaci glykemie. Tento jev významně predisponuje k T2DM.
d) M4 receptory
M4 receptory nejsou zatím zcela prozkoumány. Mechanismus jejich účinku je obdobný jako u M2 receptorů (Gi-protein snížením koncentrace cAMP aktivuje kanály pro K+). Nacházejí se zejména ve striátu, kde působí jako regulační autoreceptory cholinergních neuronů (stejný efekt mají M2 v hipokampu a jiných korových oblastech).
Ve striátu tlumí efekt D1 receptorů s excitačním efektem, jejichž prostřednictvím dopamin zvyšuje aktivitu extrapyramidového motorického systému. Fyziologické implikace této interakce nejsou ještě zcela známy.
e) M5 receptory
M5 receptory jsou stejně jako M1 a M3 spřaženy s Gq-proteinem. Většina výzkumů M5 receptorů, které byly k dnešnímu dni provedeny, proběhla in vitro.
2) Nikotinové receptory
Jde o ionotropní receptory řízené ligandem. Jedná se v podstatě o kationtové kanály propustné pro Na+ a K+, v případě některých podtypů i pro Ca2+. Jejich základní dělení rozlišuje muskulární typ (NM receptor) přítomný hlavně na nervosvalové ploténce a neuronální typ (NN receptor), který nacházíme ve všech vegetativních gangliích na membráně postsynaptického terminálu. V CNS fungují nikotinové receptory typu NN jako heteroreceptory pro jiné systémy (GABAergní, serotoninergní, glutamatinergní a dopaminergní). Jelikož jsou propustné pro Ca2+, zvyšují množství uvolněného neurotransmiteru.
Cholinergní neurony se nacházejí zejména v nc. basalis Meynerti a jádrech septa.
Tyto neurony projikují do kůry a hipokampální formace. Podílejí se na aktivaci některých korových oblastí a konsolidaci krátkodobé paměti. Cholinergní neurony nc. basalis a středního septa jsou poškozeny při Alzheimerově chorobě.
Další skupina cholinergních neuronů se nachází v tegmentu mozkového kmene a vysílají své axony do mozečku, hypotalamu a nižších oddílů CNS. Ovlivňují bdělost, spánkový cyklus a jsou významné pro iniciaci REM fáze spánku.
Ve striatu se nacházejí cholinergní interneurony, které jsou zapojeny do okruhu bazálních ganglií a podílejí se tak na kontrole postoje, iniciaci pohybů a selekci žádaných pohybových vzorců.
Klinické korelace:
Pro lékařství mají velký význam inhibitory acetylcholinesterázy. Některé z nich jsou vysoce toxické látky, jiné mohou sloužit jako léčiva některých závažných onemocnění.
Mezi velmi účinné inhibitory acetylcholinesterázy patří organofosfáty. Tyto sloučeniny tvoří kovalentní vazbu s OH skupinou serinu v molekule acetylcholiesterázy. Vazba je tak silná, že vydrží několik týdnů. Organofosfáty se nejčastěji využívají jako insekticidy, kdy mohou způsobovat náhodné otravy (symptomy jsou způsobeny nadbytkem acetylcholinu v synaptické štěrbině). Existují i vysoce toxické organofosfáty (nervově paralytické plyny), např. sarin, tabun či VX.
Inhibitory acetylcholinesterázy se využívají v léčbě takových onemocnění, jako je Alzheimerova choroba či myashenia gravis.
_
Katecholaminy jako neurotransmitery
Katecholaminy (noradrenalin, adrenalin a dopamin) jsou významnými neurotransmitery CNS, přičemž funkce každého z nich je poněkud odlišná. V této podkapitole detailně probereme zejména působení noradrenergního a dopaminerního systému.
Nejdříve se zaměříme na proces syntézy a inaktivace katecholaminů v CNS.
Syntéza a inaktivace katecholaminů
Mezi katecholaminy řadíme tři sloučeniny – adrenalin, noradrenalin (v anglosaské literatuře epinefrin a norepinefrin) a dopamin. Všechny jsou odvozeny od aromatické aminokyseliny L-tyrosinu (popř. L-fenylalaninu). Základ jejich struktury tvoří pyrokatechol (angl. catechol) – benzen se dvěma hydroxylovými skupinami, na který je navázán postranní řetězec s aminoskupinou.
Konverze tyrosinu na adrenalin zahrnuje tyto kroky:
1) Hydroxylace L-tyrosinu
2) Dekarboxylace L-dopy
3) Hydroxylace dopaminu
4) Methylace noradrenalinu
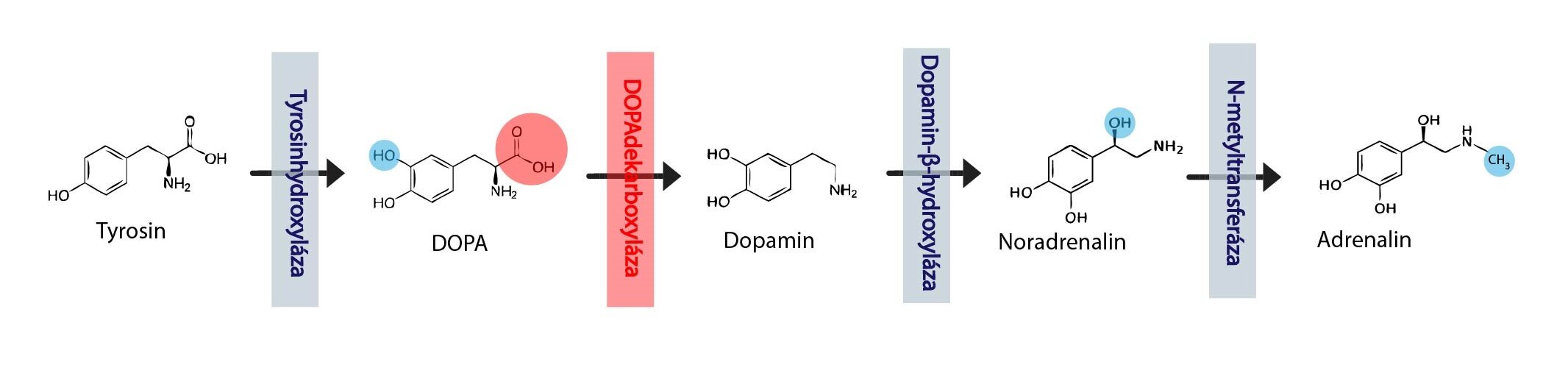
1) Hydroxylace L-tyrosinu
Při prvním kroku dochází k přeměně L-tyrosinu na L-dihydroxyfenylalanin (L-dopa). Reakci katalyzuje tyrosinhydroxyláza, která je současně rate-limiting enzymem syntézy katecholaminů. Hlavním regulačním mechanismem je feedback inhibice katecholaminy.
2) Dekarboxylace L-dopy
Při druhém kroku se L-dopa dekarboxyluje na biogenní amin 3,4-dihydroxyfenylethylamin – dopamin. Reakci katalyzuje DOPAdekarboxyláza (tento enzym dekarboxyluje všechny aromatické aminokyseliny, a proto se mu také říká dekarboxyláza aromatických aminokyselin, viz syntéza serotoninu).
Klinické korelace:
Alfa-metyldopa, která se strukturně podobá L-dopě, je kompetitivní inhibitor DOPAdekarboxylázy. Využívá se při léčbě vysokého krevního tlaku v těhotenství.
Nasyntetizované katecholaminy nedovedou překročit hemotoencefalickou bariéru (HEB), a tvoří se proto přímo v buňkách CNS. Parkinsonova choroba je doprovázena nedostatečnou tvorbou dopaminu v buňkách CNS. Při její léčbě může pomoci L-dopa, jež HEB přestupuje a posiluje tvorbu dopaminu v CNS.
3) Hydroxylace dopaminu
Třetím krokem je hydroxylace postranního řetězce dopaminu – vzniká noradrenalin. Reakci katalyzuje dopamin-β-hydroxyláza.
4) Methylace noradrenalinu
Závěrečným krokem je methylace aminosykupiny noradrenalinu za vzniku adrenalinu. Reakci katalyzuje N-methyltransferáza (fenylethanolamin-N-methyltransferáza – PNMT), kofaktorem – donorem methylu je S-adenosylmethionin (SAM).
Ukončení neurotransmise je podmíněno reuptakem katecholaminů a jejich následnou intracelulární inaktivací. Uvedeme si dva enzymy, jež ji katalyzují:
1) Katechol-o-methyl transferáza (COMT)
2) Monoaminooxidáza (MAO)
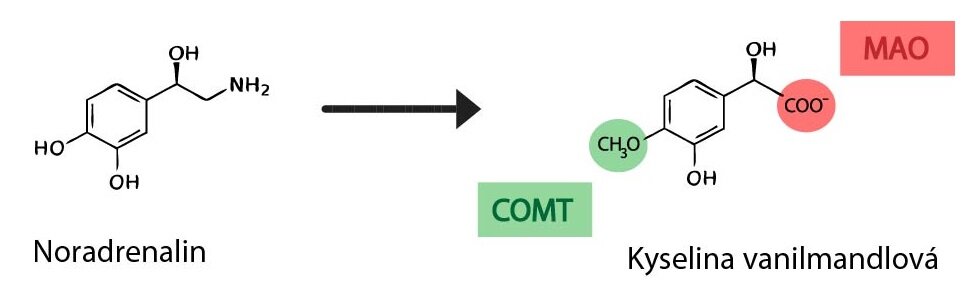
1) Katechol-o-methyl transferáza (COMT)
Katalyzuje methylaci hydroxylové skupiny katecholaminů, dárcem methylu je S-adenosylmethionin. COMT katalyzuje například tyto reakce:
Noradrenalin → normetanefrin
Adrenalin → metanefrin
Dopamin → 3-methoxytyramin
2) Monoaminooxidáza (MAO)
Katalyzuje oxidativní deaminaci katecholaminů (obecně monoaminů), kdy odstraněná aminoskupina je nahrazena aldehydovou skupinou. Ta je následně oxidována aldehyddehydrogenázou na karboxylovou skupinu. MAO (společně s aldehyddehydrogenázou) katalyzuje například tyto reakce:
Metanefrin či normetanefrin → vanilmandlová kyselina (VMA)
3-methoxytyramin → homovanilová kyselina (HVA)
MAO je mitochondriální enzym vázaný ve vnější mitochondriální membráně. MAO patří mezi flavoproteiny – obsahuje kovalentně vázaný kofaktor FAD. U lidí se vyskytují dva typy MAO: MAO-A a MAO-B. Oba dva typy se nalézají v nervové tkáni, MAO-A navíc v játrech, GIT a placentě a MAO-B navíc v krevních destičkách.
Klinické korelace:
Inhibitory MAO (IMAO, např. moklobemid a selegilin) se používají pro léčbu deprese či Parkinsonovy choroby.
Činností COMT a MAO se tvoří významný degradační produkt katecholaminů, kyselina vanilmandlová. Ta se vylučuje převážně močí, kde ji můžeme stanovit k určení produkce katecholaminů v těle. Nejčastěji se využívá při podezření na nádor dřeně nadledvin – feochromocytom.
Nyní se zaměříme na jednotlivé katecholaminergní systémy: noradrenergní a dopaminergní.
Noradrenergní systém
Noradrenalin je neurotransmiter, který v CNS působí zejména jako regulátor aktivity ostatních projekčních systémů a některých funkcí. Pokud se zvýší aktivita noradrenergních drah, dojde ke snížení nebo zvýšení excitability cílových míst v závislosti na přítomných receptorech a dalších mechanismech, které ještě nejsou zcela známy. Je nutné si uvědomit, že noradrenergní dráhy mohou takto modulovat jak excitační funkci glutamátu, tak inhibiční funkci GABA.
Účinek noradrenalinu je dán přítomností určitého typu receptoru na membráně cílové buňky. V CNS nacházíme čtyři podtypy adrenergních receptorů: α1, α2, β1 a β2.
1) α1-receptor
Nachází se na postsynaptických neuronech a zprostředkovává excitační efekt noradrenalinu.
2) α2-receptor
Nachází se obvykle na presynaptickém terminálu a má inhibiční účinky. Jeho význam spočívá v presynaptické inhibici uvolňování noradrenalinu, což je negativní zpětnovazebný regulační mechanismus.
3) β1-receptor
Neuronální receptor s excitačním efektem.
4) β2-receptor
Podtyp receptoru, který je v CNS lokalizován na buňkách glie. Poslední výzkumy ukazují, že β2-receptor se podílí na integraci dvou řídících supersystémů, a to nervového a imunitního. Přesná fyziologická role tohoto receptoru na integraci je intenzivním předmětem zkoumání.
Noradrenergní neurony se nacházejí v mozkovém kmeni, a to zejména v locus coeruleus (tzv. skupina A6), tegmentu a retikulární formaci medully a pontu (skupiny A1, A2, A5, A7).
1) Locus coeruleus
Axonální projekce neuronů z locus coeruleus vstupují doslova do všech oddílů CNS. Úkolem tohoto systému je modulovat excitabilitu ostatních projekčních systémů a přímo regulovat pozornost, bdělost, spánkový cyklus a odpověď na různé stresory. Projekce do asociačních oblastí kůry ovlivňují také emoční chování.
2) Skupiny tegmenta a retikulární formace
Neurony těchto skupin projikují do míchy, kmene, hypotalamu a limbického systému, a to zejména do těch oblastí, které se přímo podílejí na řízení viscerálních funkcí.
V míše pak regulují excitabilitu motoneuronů předního rohu.
V retikulární formaci je i menší množství neuronů, které využívají jako neurotransmiter adrenalin a jejichž aferenty jsou totožné s těmi z locus coeruleus. Má se za to, že spolupracují s noradrenergními neurony při řízení poplašné reakce a adaptaci na stresor.
Klinická korelace:
Katecholaminová hypotéza vysvětluje vznik afektivních poruch, zejména pak deprese, primárně jako hypoaktivitu noradrenergních projekcí z locus coeruleus do některých oblastí kůry (současně je ovšem přítomná i porucha serotoninergního systému).
Dopaminergní systém
Dopamin hraje v CNS významnou úlohu při řízení motorických funkcí, iniciaci různých vzorců chování a při modulaci aktivity viscerálních funkcí.
V současné době je známo pět dopaminových receptorů, tzv. D1-5, které jsou spřaženy s G-proteinem ovlivňujícím aktivitu adenylátcyklázy. Dělí se do dvou skupin dle toho, zda adenylátcyklázu stimulují prostřednictvím GS (D1, D5), nebo inhibují prostřednictvím GI (D2, D3, D4). Jednotlivé struktury CNS se liší denzitou a zastoupením jednotlivých receptorů. Např. motorické korové oblasti jsou bohaté na D2, na rozdíl od limbického systému, kde převládají receptory D3 a D4. Adrenalin a noradrenalin jsou parciální agonisté většiny dopaminových receptorů.
Neurony dopaminergního systému se nacházejí v mezencefalu a hypotalamu. Funkčně rozlišujeme několik projekcí dopaminergního systému, které jsou některými autory považovány za diskrétní systémy.
1) Nigrostriatální projekce
Neurony ze substantia nigra pars compacta vysílají své axony převážně do striata, globus pallidus a putamen. Jsou významné svým zapojením do okruhu bazálních ganglií. Podílejí se na plánování a exekuci korových aktivit, s nejsilnějším vlivem na motorickou kůru.
Hypofunkce této projekce se projeví klidovým třesem, bradykinezí, svalovou ztuhlostí a posturálním deficitem. Na poškození dopaminergních neuronů v pars compacta se mohou podílet idiopatické degenerativní změny (Parkinsonova choroba) nebo aterosklerotické zúžení lumina cév (parkinsonismus).
2) Mezolimbické projekce
Dopaminergní neurony z ventrálního tegmenta a mezencefalické retikulární formace vysílají mezolimbické projekce do nc. accumbens, amygdaly a hipokampu. Podílejí se na motivaci, odměně a trestu, pudovém chování a vzniku závislostí. Jejich zvýšená aktivita vede k exekuci stereotypních vzorců chování, opakujících se pohybů apod.
3) Mezokortikální projekce
Některé neurony ventralního tegmenta a mezencefalické retikulární formace vysílají také projekce do prefrontální oblasti kůry. Ovlivňují plánování a exekuci aktivit frontálního kortexu (což se projeví např. udržením pozornosti) a přivádějí významné podněty pro iniciaci chování.
Mezolimbické a mezokortikální projekce spolu úzce souvisí a jsou cílem neuroleptik a antipsychotik při farmakologické terapii (např. schizofrenie, obsedantně kompulzivní poruchy, hyperaktivní poruchy pozornosti (ADHD) a Tourettova syndromu).
4) Tuberoinfundibulární projekce
Tlumí sekreci prolaktinu.
5) Intrahypotalamická projekce
Podílí se na modulaci různých viscerálních funkcí.
Klinická korelace:
Dnes se má za to, že schizofrenie je alespoň zčásti vyvolána excesivní aktivitou mezolimbického systému a útlumem systému mezokortikálního.
_
Serotoninergní systém
Serotoninergní systém zastává v CNS podobnou úlohu jako systém noradrenergní. Reguluje celou řadu funkcí a moduluje aktivitu ostatních projekčních systémů. Velmi často dokonce inervuje i zcela totožné struktury a jejich noradrenergní aferenty funkčně doplňuje.
Dosud bylo objeveno sedm podtypů serotoninergních receptorů, tzv. 5-HT1-7R, z nichž některé jsou považovány za excitační a jiné za inhibiční. Efekt serotoninu je tedy velmi závislý na lokalitě, respektive receptorech, které jsou exprimovány neurony dané struktury.
Serotoninergní neurony se nacházejí v rafeálním jádře retikulární formace, které lze funkčně rozdělit na část rostrální s ascendentní eferentací a část kaudální s descendentní eferentací.
1) Ascendentní serotoninergní systém (nc. raphe dorsalis, nc. raphe pontis centralis superior)
Vysílají projekce do kůry, některých struktur limbického systému (hipokampus, amygdala), bazálních ganglií, celé řady jader a areí hypothalamu a některých jader thalamu.
Tento systém se podílí na regulaci emočního chování a hypothalamických funkcí (zejména viscerálních) a řídí spánkový cyklus. Dále moduluje zpracování informací v senzorických oblastech (např. ve vizuální kůře).
2) Descendentní serotoninergní systém (nc. raphe pontis, nc. raphe magnus, nc. raphe obscurus)
Projekce této části rafeálního jádra inervují zejména kaudálněji položené části CNS: mozkový kmen, mozeček a míchu.
Kaudálně uložené neurony modulují vnímání bolesti projekcemi do zadního rohu míšního, přičemž zvýšená aktivita serotoninergního systému posiluje efekt anestetik a je zcela nezbytná pro působení opiátů. Jejich projekce dále modulují excitabilitu pregangliových neuronů autonomního nervového systému a zvyšují excitabilitu motoneuronů v předních rozích míšních.
Klinická korelace:
Léky zvané SSRI (specifické inhibitory zpětného vychytávání serotoninu) zvyšují aktivitu serotoninergního systému a jsou velmi účinné při léčbě pacientů s unipolární depresí. Zároveň ale zmenšují libido a způsobují poruchy příjmu potravy, obvykle charakteru hyperfagie.
Syntéza a inaktivace serotoninu
Serotonin je odvozen od L-tryptofanu, jedná se o 5-hydroxytryptamin (5-HT). Jeho tvorba z tryptofanu probíhá ve dvou krocích:
1) Hydroxylace tryptofanu
2) Dekarboxylace
1) Hydroxylace tryptofanu
Prvním krokem je hydroxylace tryptofanu v poloze 5, čímž vzniká 5-hydroxytryptofan. Reakci katalyzuje tryptofanhydroxyláza, která je současně rate-limiting enzymem syntézy serotoninu.
2) Dekarboxylace
Dekarboxylací 5-hydroxytryptofanu vzniká 5-hydroxytryptamin neboli serotonin, který tedy patří mezi biogenní aminy (stejně jako všechny katecholaminy). Reakci katalyzuje 5-hydroxytryptofandekarboxyláza (dekarboxyláza aromatických aminokyselin, viz syntéza katecholaminů).
Ukončení neurotransmise je podmíněno reuptakem a následnou intracelulární inaktivací serotoninu. Inaktivaci serotoninu katalyzuje dvojice enzymů – monoaminooxidáza (MAO) a aldehyddehydrogenáza (viz inaktivace katecholaminů).
Působením těchto dvou enzymů vzniká 5-hydroxyindolacetát (kyselina 5-hydroxyindoloctová – 5-HIAA), který se z těla vylučuje převážně močí (jako konjugát s kyselinou glukuronovou). Jeho stanovení se využívá k diagnostice nádorů produkujících serotonin – neuroendokrinní tumory (dříve karcinoid).
U serotoninu existuje ještě jedna boční metabolická dráha, při které se ze serotoninu stane hormon melatonin. Nejdříve navážeme na aminoskupinu serotoninu acetyl – vzniká N-acetylserotonin. Následně se na hydroxylovou skupinu naváže methyl – vzniká melatonin.
Historická korelace:
Název serotonin se odvozuje od jeho objevu. Serotonin totiž poprvé popsali jako látku krevního séra účinkující na tonus hladkého svalstva.
_
Glycinový systém
Glycin je neurotransmiter, který vykazuje významnou inhibiční aktivitu. Ta je v CNS zprostředkována přes ionotropní receptor spřažený s Cl– kanálem (vyvolá hyperpolarizaci membrány). Ve velké koncentraci jej nacházíme hlavně v šedé hmotě míšní, kde je hlavním neurotransmiterem inhibičních interneuronů.
Mimo své inhibiční působení je i alosterickým modulátorem aktivity NMDARs. Má tedy i excitační schopnost ve smyslu facilitace aktivity glutaminergního systému.
Klinické korelace:
Strychnin je inhibitor glycinového receptoru. Tím, že inhibuje inhibiční interneurony v šedé hmotě míšní, umožňuje nekoordinované rozšíření podráždění. Vznikají rychle se šířící záchvaty křečí. Strychnin má velmi výraznou hořkou chuť, patří mezi nejvíce hořké látky vůbec.
_
Histaminergní systém
Histamin zastává v těle celou řadu rozličných funkcí, jako neurotransmiter se podílí zejména na regulaci spánku, bdělosti a sekreci hormonů z hypotalamohypofyzárního systému.
Strukturou se jedná o biogenní amin vznikající dekarboxylací histidinu (enzym histidindekarboxyláza). Uvolněný histamin je rychle degradován diaminoxidázou.
O účincích histaminu rozhodují receptory H1, H2, H3. Ty se vyskytují jak v CNS, tak v periferních tkáních.
1) Receptory H1
Jsou exprimovány ve velké denzitě v hypotalamu a mammilárních tělíscích, jejich vliv je exitační. Aktivace H1R vede k snížení permeability membrány pro K+, což způsobuje depolarizaci. Většina antihistaminik vyvolává únavu právě antagonizujícím působením na H1 receptory.
2) Receptory H2
Jsou to metabotropní receptory spřažené s GS-proteinem, nacházejí se na neuronech mnoha oblastí kůry, ale i na gliích a endotelu mozkových cévních kapilár.
3) Receptory H3
Nacházejí se na presynaptických terminálech a jako autoreceptory tlumí další vyplavování histaminu. Lze je také nalézt na noradrenergních, dopaminergních a cholinergních (aj.) neuronech, kde jako heteroreceptory tlumí vyplavení transmiteru příslušného systému.
_
Purinergní neurotransmitery
Některé puriny fungují jako neurotransmitery – např. ATP a ostatní trifosfáty, AMP či adenosin.
Purinergní receptory neboli purinoceptory jsou membránové receptory přítomné na většině buněk lidského těla. Plní nepřeberné množství funkcí – od indukce proliferace a spoluřízení migrace buněk neurální lišty v embryonálním období, přes navození apoptózy a sekrece cytokinů.
Základní dělení rozlišuje tři třídy purinergních receptorů:
1) P1 receptory
2) P2X receptory
3) P2Y receptory
1) a 3) P1 receptor a P2Y receptor
Jde o receptory spřažené s G-proteinem. Nacházíme je zejména v srdci, ledvinách, tukové tkáni a mozku. Vzájemně se liší svým hlavním agonistou, tím je u P1 adenosin a u P2Y ATP. Navíc receptor P2Y hraje významnou úlohu v patofyziologii chronické bolesti.
2) P2X receptor
Jedná se o receptor spřažený s neselektivním kationtovým kanálem (ligand-gated ion channel), jehož hlavním agonistou je ATP. P2X receptor je zodpovědný za udržení postsynaptických excitačních odpovědí. Nachází se zejména v centrálním a periferním nervovém systému, a to jak na neuronech a gliích, tak i na cévách zásobujících nervovou tkáň. Toto rozložení receptorů je pravděpodobně funkčně významné během vývoje nervové soustavy a během chorobných stavů (záněty, nádory), neboť integruje a koordinuje odpověď jednotlivých nervových a cévních elementů.
Nedávné studie potvrdily roli tohoto receptoru v rozvoji neuropatické bolesti.
_
Peptidy
Peptidové neurotransmitery jsou syntetizovány – jako každý jiný protein – na ribosomech. Jejich tvorba probíhá v těle neuronu, odkud jsou transportovány axonálním přenosem do nervového zakončení.
Obecně se dá říci, že na začátku se vytvoří velká mateřská molekula, jež je následně rozstříhána na menší peptidy, které jsou aktivními neurotransmitery. Na tomto místě si uvedeme jeden příklad mateřské molekuly – POMC (proopiomelanokortin, 241 AK). Ta se štěpí na několik dceřiných neurotransmiterů:
1) β-endorfin
2) MSH (melanocyty stimulující hormon)
3) ACTH (adrenokortikotropní hormon)
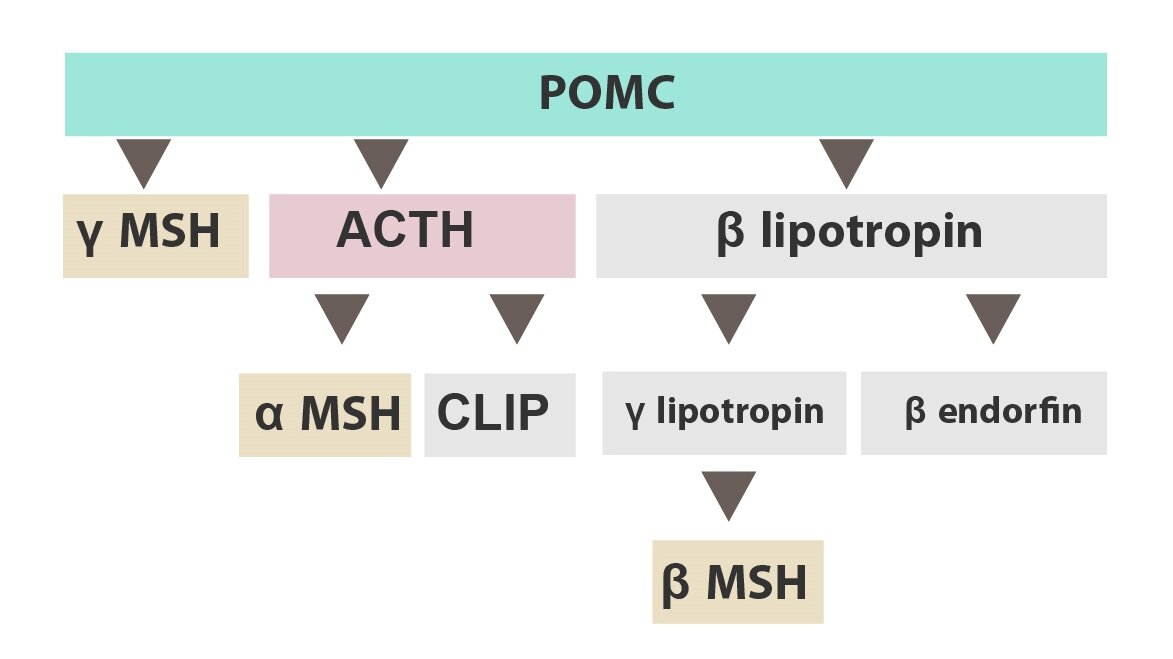
Z proopiomelanokortinu mohou vznikat i další molekuly. Tři výše uvedené zde uvádíme, protože pomáhají vysvětlit název mateřské molekuly.
Skupina peptidových neurotransmiterů je velmi široká. Uvedeme si zde proto jen několik příkladů:
1) Opioidní substance (váží se na opioidní receptory): enkefaliny, endorfiny, dynorfin
2) Substance P
3) Neuropeptid Y
4) Somatostatin
5) Cholecystokinin
_
Endogenní kanabonoidy
Jejich zástupcem je například anandamid (arachidonoylethanolamid, AEA) – derivát kyseliny arachidonové a etanolaminu. Syntéza a degradace anandamidu je poměrně složitá a nebudeme ji zde popisovat, zájemce odkazujeme na doporučenou literaturu.
Anandamid působí v nervovém a imunitním systému prostřednictvím kanabinoidových receptorů CB1 a CB2.
1) CB1 receptory
Nacházejí se ve velké denzitě v centrálním nervovém systému. Jsou spřaženy s G-proteinem a jsou exprimovány zejména presynapticky na GABAergních neuronech. Při jejich aktivaci dochází k snížení množství kvant GABA, která se uvolní do synaptické štěrbiny. Tento jev se nazývá depolarizací indukovaná suprese inhibice (DSI).
Agonisté CB1 receptorů mají orexigenní účinky a stimulují centra odměny.
2) CB2 receptory
Jsou přítomny na neuronech periferního nervového systému, hematopoetických a imunokompetentních buňkách. Hrají významnou roli v antinocicepci a rozvoji imunitní odpovědi.
Historická korelace:
Strukturu anandamidu popsal v roce 1992 český analytický chemik Lumír Ondřej Hanus a americký farmakolog William Anthony Devane na Hebrejské univerzitě v Jeruzalémě. Jeho název je odvozen ze sanskrtského ananda (blaženost, dokonalé štěstí).
Autoři podkapitoly: Patrik Maďa a Josef Fontana
![]()