Náplň podkapitoly:
1. Úvod do motorických funkcí
2. Reflexy a svalové receptory
3. Centrální řízení motoriky
_
Úvod do motorických funkcí
Spolu s tím, jak náš centrální nervový systém sbírá a vyhodnocuje senzorické informace, musí být vybaven systémy, které mu umožní na daný podnět patřičně zareagovat. Nejběžnější reakci na většinu stimulů představuje motorická odpověď.
Tento trend je společný všem živočichům napříč taxony. Proto se nezmar při mechanickém podráždění stáhne, srna při sebemenší známce nebezpečí uteče, zatímco hladový predátor se vydá hledat potravu.
Reflexy a svalové receptory
Míšní reflexy
V této části se budeme nejdříve zabývat míšními reflexy. Reflex je základní funkční jednotkou motoriky. Jejich morfologickým podkladem jsou specializované neuronální okruhy, které řídí funkci svalů tak, aby daly vznik účinným pohybům. Bez těchto okruhů by nemohly být provedeny ani ty nejjednodušší pohyby.
Role vyšších motorických center totiž spočívá převážně v tom, že při patřičném stimulu aktivují míšní reflexní okruhy, a to v patřičném pořadí a s přiměřenou intenzitou. Po přerušení míchy nelze chodit, ani když je plně zachována funkce vyšších motorických center, poněvadž motorické oblasti kůry nemají přístup k aktivaci neuronálního okruhu pro chůzi.
Reflexní oblouk
Základní uspořádání míšního reflexu nazýváme reflexní oblouk. Ten je složen z následujících částí:
1) Senzor
2) Aferentní dráha (dostředivá)
3) Centrum reflexu
4) Eferentní dráha (odstředivá)
5) Efektor
Obecný princip činnosti reflexního oblouku je relativně prostý. Senzor zachytí adekvátní vjem, jejž aferentní dráha vede do centra reflexu, kde je vyhodnocen a integrován s dalšími adekvátními podněty. Tím vznikne informace odeslaná přes eferentní dráhu k efektoru. Ten nakonec zprostředkuje patřičnou motorickou odpověď.
Senzor
Za senzor reflexu můžeme považovat jakýkoliv receptorový aparát, jehož podráždění vyvolá reflexní odpověď. Může tedy například jít o volná nervová zakončení registrující bolest nebo o Golgiho tělíska v úponech svalů.
Aferentní dráha
Aferentními dráhami rozumíme převážně vlákna, která pocházejí z neuronů spinálních ganglií. Táhnou se od příslušného receptoru a do CNS vstupují zadními rohy míšními. V míše se okamžitě dělí na dvě kolaterály:
1) Kolaterála končí na tělech a dendritech neuronů v šedé hmotě příslušného míšního segmentu – vyvolává místní reflexy
2) Kolaterála postupuje dále v CNS a končí ve vzdálenějších míšních segmentech nebo neuronech mozkového kmene či kůry (převážně v senzorických oblastech)
Centrum reflexu
Pokud prozatím zůstaneme u popisu jednodušších reflexů, lze tvrdit, že centrum reflexu spočívá v šedé hmotě míšní. Zde se nachází několik miliónů neuronů, které přijímají informace ze senzorických systémů. Pro motorické reflexy jsou pak obzvlášť významné motorické neurony předního rohu a interneurony.
1) Motorické neurony
Jedná se o velké neurony, jejichž axony jsou v reflexologické terminologii nazývány eferentními vlákny. Opět je můžeme rozdělit do několika skupin, z nichž nejvýznamnější jsou alfamotoneurony a gammamotoneurony.
a) Alfamotoneurony
Alfamotoneurony přímo odpovídají za aktivaci efektoru. Dávají vznik objemným myelinizovaným axonům, jež vedou vzruchy velmi rychle. Na konci svého průběhu se větví a končí tzv. nervosvalovou ploténkou (neuromuskulární junkcí) na svalových vláknech. Jeden motoneuron inervuje obvykle více svalových vláken, ta se při jeho aktivaci stáhnou současně.
Všechna svalová vlákna inervovaná jedním alfamotoneuronem se označují pojmem motorická jednotka. Motorická jednotka představuje ve fyziologii významný pojem. Síla stahu daného svalu totiž závisí pouze na počtu aktivovaných motorických jednotek. Jejich počet udává citlivost motorického řízení pro daný sval. Tak například m. quadriceps femoris adaptovaný na hrubé pohyby se skládá z velkých motorických jednotek, kde jeden alfamotoneuron inervuje stovky svalových vláken. Naopak motorické jednoty okohybných svalů jsou malé a alfamotoneurony obvykle inervují jednotlivá svalová vlákna, čímž dovolují přesné řízení na zlomky milimetrů.
b) Gammamotoneurony
Axony těchto motorických neuronů, významně menších než alfamotoneurony, končí na speciálních svalových vláknech, tzv. intrafuzálních vláknech tvořících speciální regulační aparát, svalové vřeténko, jež se účastní kontroly svalového tonu. Podrobněji se o něm zmíníme dále v textu.
2) Interneurony
Interneurony se nacházejí difúzně v celém míšním segmentu. Je jich přibližně třicetkrát více než motorických neuronů. Jsou hojně propojeny jak mezi sebou navzájem, tak s motoneurony, čímž tvoří složitou síť, která odpovídá za integrační funkce míchy. Takovouto síť funkčně spřízněných neuronů nazýváme neuronálním poolem a motiv, dle kterého jsou propojeny, neuronálním okruhem. Tyto okruhy nabývají určitých vlastností: mohou být divergující, konvergující, reverbující, se sériovým zpracováním a s paralelním zpracováním.
a) Divergence
Divergencí chápeme šíření vzruchu z jednoho neuronu na celou řadu dalších. Jinak řečeno, informace je předána z jednoho neuronu alespoň na dva další, které s prvním neuronem sdílejí synapse.
b) Konvergence
Konvergencí nazýváme situaci, při které jeden neuron přijímá informaci alespoň ze dvou různých neuronů současně.
c) Reverberace
Reverberace označuje situaci, kdy jedna informace opakovaně krouží neuronálním okruhem. Toho je obvykle dosaženo buď kolaterálou neuronu, která je napojena na neuron předchozí, nebo několika sériově zapojenými neurony tvořícími samotný uzavřený okruh (tzv. repetitivní okruh).
d) Sériové zpracování informace
Situaci, kdy jeden neuron předá danou informaci právě jednomu následujícímu atd., označujeme jako sériové zpracování informace. Neurony v něm tvoří vzájemně propojený řetězec.
e) Paralelní zpracování informace
Paralelní zpracování probíhá ve chvíli, kdy je stejná informace vedena alespoň dvěma neurony současně a vždy jí musí předcházet divergence. Tím je docíleno toho, že na jeden podnět aktivuje více efektorů. Např. došlápnutí na ostrý objekt (bolestivý podnět) aktivuje flexi v poraněné končetině a extenzi v kontralaterální.
Většina vstupů z vyšších motorických etáží CNS je zakončena právě na příslušném neuronálním poolu. Stejně tak existuje pouze několik málo reflexních okruhů, které končí na motoneuronu. Jsou však velmi významné, a proto je budeme dále v textu detailně diskutovat.
Eferentní dráha
Eferentní dráha vystupuje z předního rohu míšního a sestává z axonů motoneuronů zakončených na příslušném efektoru.
Efektor
Je to orgán složený z dráždivých buněk, jehož funkci reguluje daný reflexní oblouk. V rámci motoriky se jedná převážně o příčně pruhovanou svalovinu. Je však nutné si uvědomit, že existují i viscerální reflexy, jejichž efektorem může být hladká svalovina nebo nejrůznější žlázy.
Svalové receptory
Než postoupíme dále v popisu základních principů řízení motoriky, je dobré si připomenout, že pro správnou funkci pohybového aparátu má neustálý zpětnovazebný monitoring aktuálního stavu svalů přinejmenším stejnou důležitost jako samotná aktivace motorických jednotek. K získávání informací, jako je změna délky nebo tenze v daném svalu, slouží dva vysoce specializované senzorické receptory:
1) Svalové vřeténko
2) Golgiho šlachové tělísko
Svalové vřeténko
Poměrně drobná svalová vřeténka se skládají z modifikovaných svalových vláken, tzv. intrafuzálních vláken, která mají zachované kontraktilní vlastnosti, ale současně slouží jako receptory protažení svalu nebo změny rychlosti protažení. Celé svalové vřeténko je tvořeno obvykle pouze 4 až 12 intrafuzálními vlákny. Svalová vřeténka jsou připojena paralelně k vlastnímu svalu na glykokalyx myocytů. Vlákna vlastního svalu označujeme jako extrafuzální vlákna.
Intrafuzální vlákna se liší od extrafuzálních tím, že jejich střední část je chudá na kontraktilní elementy a slouží pouze jako receptorová oblast. Vjem je zachycen vždy, když dojde k protažení střední porce intrafuzálního vlákna. Vlastní kontrakce vlákna tak pramení pouze v okrajových částech vlákna, jež jsou inervována gammamotoneuronem.
Díky tomuto uspořádání je svalové vřenténko aktivováno ve dvou situacích:
1) Protažení extrafuzálních vláken, na které je připojeno – tím se protáhne i receptorová část vřeténka
2) Zvýšenou aktivitou gammamotoneuronu, neboť se kontrahují pouze okrajové části vlákna a receptorová porce se tak protáhne
Tyto změny snímá dvojí uspořádání senzorických nervových zakončení:
1) Primární zakončení (annulospirální vlákna)
2) Sekundární zakončení
Jak již název napovídá, annulospirální nervová zakončení se obtáčejí kolem intrafuzálního vlákna a nacházíme je asi ve středu jeho délky. Sekundární nervová zakončení mají spíše rozvětvenou keříkovitou strukturu a jsou umístěna na okrajových částech vřeténka.
Dvojí uspořádání vykazují i samotná intrafuzální vlákna. Rozlišujeme:
1) Vlákna s řetězcem jader
2) Vlákna s jaderným vakem
Ta se vzájemně liší uspořádáním jader v centrální receptorové části. Vlákna s řetězcem jader mají jádra uspořádána za sebou, čímž vzniká vlákno, jehož průměr se v průběhu příliš nemění. U vláken s jaderným vakem jsou jádra nakupena ve střední části, což vede k jejich výraznému vyklenutí.
Tento morfologický jev je funkčně velice významný. Pokud se totiž bude protahovat vlákno s jaderným řetězcem, dojde k jeho významnému zúžení v celé jeho délce a aktivuje jak primární, tak sekundární nervová zakončení. Naopak vlákna s jaderným vakem mají v centrální porci takový dostatek „nadbytečné plochy membrány”, aby nedošlo k významnému zúžení na koncích intrafuzálního vlákna. Dojde tedy ke změně průměru v oblasti jaderného vaku (kde se nacházejí annulospirální zakončení), nikoli v krajních pozicích, které jsou snímány sekundárními nervovými zakončeními. Vlákna s jaderným vakem tedy neaktivují sekundární nervová zakončení, čímž umožňují existenci dvou typů odpovědi – statické a dynamické.
Statická odpověď
Statická odpověď nastává pokud jsou simultánně aktivována jak primární, tak i sekundární nervová zakončení. Tento jev nastává při pomalém protažení svalu a trvá i řadu minut po ukončení kontrakce, jakmile sval dosáhl konečného protažení. Statická odpověď tak v každém okamžiku informuje o aktuálním protažení svalu. Velikost protažení je přímo úměrná frekvenci výbojů jak v primárních, tak v sekundárních nervových zakončeních.
Jinými slovy: čím více je intrafuzální vlákno v daném okamžiku protaženo, s tím vyšší frekvencí produkuje akční potenciály. Centrum reflexu (ale i vyšší motorická centra) tak získává přesnou kódovanou informaci.
Dynamická odpověď
Dynamická odpověď nastává, je-li sval protažen rychle. Při něm dojde k excesivní aktivaci primárních nervových zakončení, neboť se prudce změní zejména průměr jaderného vaku. Taková reakce svalového vřeténka, kdy naprosto převažuje aktivita v annulospirálních nervových zakončeních, se nazývá dynamická odpověď a kóduje rychlost změny protažení. Po zastavení kontrakce opět dochází k simultánní aktivitě v primárních i sekundárních vláknech, která informuje o aktuální délce svalu.
Dynamická odpověď se také vyznačuje mnohem vyšší frekvencí akčních potenciálů na primárních zakončeních než u odpovědi statické.
Regulace intenzity odpovědi
Vyšší nervová centra, zejména pak bulboretikulární oblast (která je pod vlivem mozečku, bazálních ganglií a motorického kortexu), ovlivňují citlivost svalového vřeténka změnou aktivity v gammamotoneuronech. Zvýšení frekvence výbojů gammamotoneuronů nutně vede ke kontrakci intrafuzálních vláken a tím i k jejich napnutí. Takto se zvýší citlivost centrální receptorové oblasti.
Bulboretikulární oblast je dokonce schopna zvlášť měnit citlivost dynamické a statické odpovědi. Byly objeveny dva typy gammamotoneuronů: gamma-d, které převážně inervují vlákna s jaderným vakem, čímž modulují dynamickou odpověď, a gamma-s objevujících se na vláknech s jaderným řetězcem a zasahujících spíše do statické odpovědi.
Napínací reflex
Kdykoli se sval prudce napne, nastane reflexní kontrakce. Tento fenomén, tzv. napínací reflex, je nejjednodušší manifestací funkce svalových vřetének. Jedná se o tzv. monosynaptický reflex a závisí pouze na dynamické odpovědi.
Prudké protažení svalu způsobí vysokofrekvenční aktivitu v annulospirálních vláknech. Ta vstupují zadním rohem míšním do příslušného segmentu, jím prostupují a končí v ipsilaterálním předním rohu přímo na alfamotoneuronu příslušného svalu. Dojde k jeho prudké aktivaci, která vyústí ve stejně prudkou kontrakci („škubnutí”). Tento reflex tak kompenzuje prudké a nečekané změny protažení.
Představte si například situaci, kdy přenášíte krabici s knihami a někdo vám do ní nečekaně přihodí několik učebnic. V této chvíli dojde k prudkému protažení zejména m. biceps brachii, ale ve zlomcích sekundy zafunguje napínací reflex a protažení vykompenzuje. Vy tak krabici udržíte a nemusíte sbírat knihy ze země.
Důvod, proč vám bedna i s knihami zůstane v náručí i po vyhasnutí excesivní dynamické odpovědi, spočívá v tom, že po iniciální prudké kompenzaci dojde ke zvýšené aktivitě i ve statické odpovědi, která udrží zvýšenou aktivitu v alfamotoneuronech m. biceps brachii. Díky tomuto jevu někteří autoři uvádějí, že napínací reflex má dvě složky: dynamický napínací reflex a statický napínací reflex.
Statický napínací reflex tedy střídá onen dynamický s určitým zpožděním daným jednak samotným charakterem odpovědi (nižší frekvence, aktivita i v sekundárních nervových vláknech) a jednak tím, že přímo na alfamotoneuronu končí pouze axony neuronů z primárních nervových zakončení. Ta sekundární končí na některém interneuronu a jsou zpracovány příslušnými neuronálními okruhy.
Klinická korelace:
Napínací reflex má i své klinické využití v neurologii. Je jednoduchým a spolehlivým testem citlivosti reflexu a tím i správné funkce CNS. Reflex lze vyvolat poklepem na šlachu příslušného svalu, čímž dojde k prodloužení celého svalu (dynamická odpověď je fyziologicky funkční již při zkrácení intrafuzálního vlákna o více než 20 mikrometrů) a pozorujeme záškub. Takto klinicky využitelných napínacích reflexů existuje celá řada, jmenují se dle svého svalu. Za všechny uveďme např. patelární, tricipitový nebo maseterový.
Role svalových vřetének ve volní motorice
Kdykoli je zahájena exekuce nějakého pohybu, dojde k simultánní aktivaci gammamotoneuronů (tzv. koaktivace alfa-gamma). Tím se zároveň a úměrně kontrahují jak extrafuzální, tak intrafuzální vlákna. Svalové vřeténko je tedy v každém okamžiku úměrně protaženo vůči monitorovanému svalu a může plnit svoji receptorovou funkci.
Svalová vřeténka se dále účastní stabilizace polohy během motorické aktivity. Při jakékoliv motorické činnosti je pozorovatelná zvýšená aktivita v bulboretikulární oblasti. Nejde ale o běžnou alfa-gamma koaktivaci. Automaticky dochází ke zvýšení citlivosti vřetének jak svalového antagonisty, tak agonisty. Pokud se zkrátí vřeténka protilehlých svalů, dojde k významné stabilizaci kloubu v zamyšlené pozici. To je dáno tím, že citlivá vřeténka by i při malých výkyvech z této polohy vyvolala napínací reflex, který by změnu kompenzoval.
Golgiho šlachový reflex
Jedná se o další významný monosynaptický reflex. Jeho senzorem je obalené Golgiho šlachové tělísko zařazené do vazivových vláken svalového úponu. Díky tomuto zařazení do série s monitorovaným svalem nemůže získat informace o délce svalu. Veličina, kterou snímá, je svalové napětí. Stejně jako svalové vřeténko poskytuje i Golgiho šlachové tělísko dynamickou a statickou odpověď. Děje se tak na jednodušším principu spočívajícím v tom, že změny svalového napětí jsou kódovány jako vysokofrekvenční aktivita a konstantní stav je vyjádřen v nižších frekvencích impulzů.
Jeho aferentní vlákno vstupuje do CNS zadním rohem míšním a je ukončeno v předním rohu na alfamotoneuronu příslušného svalu. Na rozdíl od napínacího reflexu se ale jedná o inhibiční synapsi. Je důležité zdůraznit, že aferent jak z Golgiho šlachového reflexu, tak z napínacího reflexu vysílají kolaterálu, která končí ve vyšších etážích CNS. Motorická centra tedy mají v každém okamžiku aktuální informace o stavu všech svalů v těle.
Tento reflex chrání svalstvo, jež by mohlo být poškozeno vysokým napětím. Pokud je překonán kritický práh pro spuštění Golgiho reflexu, je během zlomku sekundy inhibován alfamotoneuron daného svalu a dojde k jeho relaxaci, tento jev se nazývá prodlužovací reakce. Další funkcí Golgiho reflexu je rovnoměrné rozdělení napětí mezi jednotlivá svalová vlákna. V každé šlaše se totiž nachází celá řada Golgiho tělísek, která tak mohou snímat i rozložení napětí v rámci jednoho svalu. Dojde-li k přetížení některých svalových vláken, Golgiho reflex inhibuje pouze alfamotoneuron příslušné motorické jednotky a tenze je v rámci svalu redistribuována.
Flexorový reflex
Flexorový reflex slouží jako příklad polysynaptického reflexu. Je též nazýván nociceptivním reflexem, neboť jeho senzorem jsou nociceptory a nejmocnějším spouštěčem právě bolest. Bolestivý stimulus je veden aferenty do zadních rohů míšních, kde i končí na přítomných interneuronech. Tím se aktivuje celý příslušný neuronální okruh, který jednak tyto informace odešle do vyšších etáží CNS, jednak ale vyvolá i lokální reakce v příslušném míšním segmentu. Po aktivaci mnoha interneuronů dojde k:
1) Aktivaci alfamotoneuronu příslušného flexoru
2) Inhibici alfamotoneuronu protilehlého extenzoru (antagonisty)
3) Propagaci do přilehlých míšních segmentů
4) Aktivaci zkříženého extenzorového reflexu
Současná inhibice svalového antagonisty brání vyvolání napínacího reflexu, který by jinak při prudké mimovolní aktivaci flexoru vznikl. Ve výsledku tedy dojde k úniku od bolestivého stimulu. Proto je flexorový reflex často charakterizován jako reflex obranný.
Jako příklad lze uvést kontakt se žhavým předmětem. Pokud si připravujeme večeři a dotkneme se rozpálené pánve, nastane mimovolní reakce, která vzdálí celou horní končetinu od bolestivého stimulu. Pokud by současně nebyl inhibován antagonista, došlo by k prudkému protažení jeho svalového vřeténka. Napínací reflex by tuto změnu vykompenzoval a ruku přitiskl zpět na pánev.
Zkřížený extenzorový reflex
Zkřížený extenzorový reflex, další z polysynaptických obranných reflexů, navazuje na flexorový reflex. Je vyvolán automaticky asi 0,5 s po reflexu flexorovém. Toto zpoždění zapříčiňuje násobné přepojování v neuronálním poolu daného segmentu. Jeho funkcí je extenze kontralaterální končetiny (oproti končetině postižené, kterou označujeme jako ipsilaterální). U lidí má význam zejména u poranění dolních končetin, lze jej však vyvolat i na končetině horní. Extenze kontralaterální končetiny zabraňuje pádu, který by nutně následoval, kdyby došlo pouze k flexi končetiny ipsilaterální.
Tak si můžeme představit, že při došlápnutí na hřebík je nejprve aktivován flexorový reflex, jenž ipsilaterální poraněnou končetinu rychlou kontrakcí flexorů zvedne z podložky. Po extenzi kontralaterální končetiny přeneseme váhu na ni. Kdyby k této extenzi nedošlo, měli bychom obě dolní končetiny flektované ve stejný okamžik a následoval by pád. S největší pravděpodobností navíc na zdroj bolestivého stimulu, v našem příkladě na onen hřebík.
Ve skutečnosti se této reakce účastní i horní končetiny, neboť jak bylo zmíněno výše, je stimulus propagován i do vzdálenějších segmentů. U horních končetin dojde také k aktivaci flexorového a zkříženého reflexu, ale stranově obráceně než u dolních končetin. To znamená, že pokud byl flexorový reflex vyvolán např. na pravé dolní končetině, nastane u pravé horní končetiny extenze a levá horní končetina bude naopak mimovolně flektována. Tato synchronní aktivace páru flexorový – zkřížený extenzorový reflex i u horních končetin je o poznání slabší než na končetinách dolnícg, kde byl aktivován senzor reflexu.
Uvedený fenomén významně přispívá k lepšímu rozložení váhy a tím k udržení rovnováhy. Evolučně se však pravděpodobně jedná o rudiment z předchozího kvadrupedického stadia.
_
Centrální řízení motoriky
Do této chvíle jsme se zmiňovali zejména o nejrůznějších reflexech, které jsou mimovolních a které tudíž z definice vůlí nelze ovlivnit. V dalším textu si přiblížíme volní motoriku.
Motorická kůra
Většina volní motoriky je iniciována v kůře. Ta jednoduše řečeno vysílá dobře naplánovanou sekvenci příkazů k neuronálním okruhům míchy, jež tak v přesném pořadí aktivuje. Existují však i výjimky, kdy samotná motorická kůra přímo aktivuje alfamotoneurony některých svalů. Obvykle se jedná o svaly s malými motorickými jednotkami, které vyžadují naprosto přesné vědomé řízení, jak to vidíme například u svalů ruky.
Motorická kůra se nachází před sulcus centralis a zaujímá asi jednu třetinu plochy frontálního laloku. Samotný motorický kortex můžeme rozdělit na tři oblasti s různými funkcemi:
1) Primární motorická oblast
2) Premotorická oblast
3) Suplementární motorická oblast
Primární motorická oblast
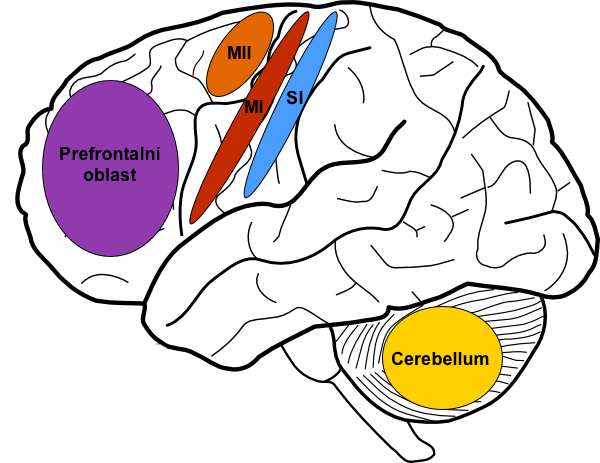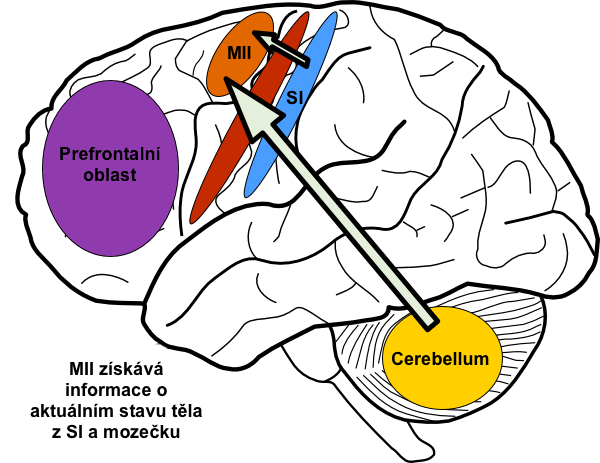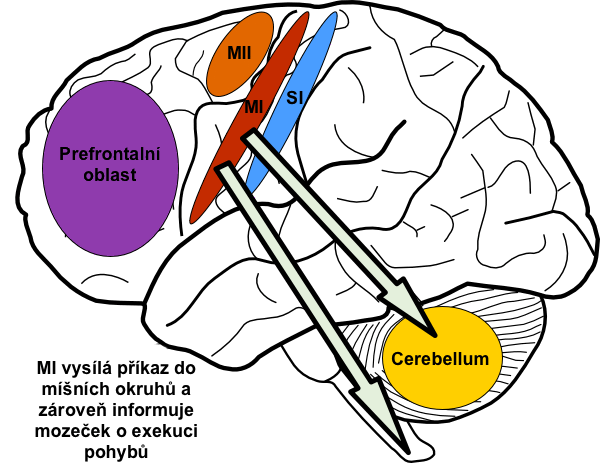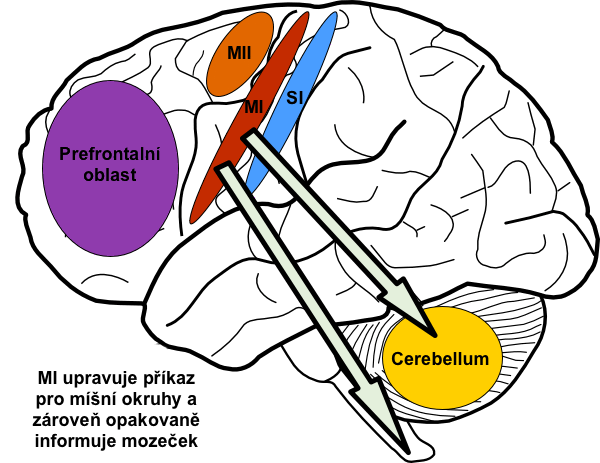
Primární motorická oblast (MI) neboli také area 4 v Brodmanově klasifikaci se nachází na gyrus praecentralis a táhne se laterálně až k fissura Sylvii. Lidem, kteří podstoupili neurochirurgické operace, byla v minulosti prováděna elektrická stimulace této oblasti. Zjistilo se, že MI je přísně somatotopicky organizována a její jednotlivé úseky můžeme přiřadit k částem těla. O této místní reprezentaci často hovoříme jako o motorickém homunkulovi, neboť na povrch g. praecentralis lze promítnout zmenšenou lidskou postavu, aby daná část těla ležela na své místní reprezentaci motorické. Tato postavička však je značně deformovaná, neboť okrsky kůry jsou proporční vzhledem k úrovni inervace, nikoli k reálné velikosti části (homunkulus má největší ruce). Tak bylo také zjištěno, že spíše než aby jeden pyramidový neuron kůry ovládal jeden sval, vetšinou zprostředkovává daný specifický pohyb, například opozice palce a ukazováku.
Premotorická oblast
Premotorická oblast se nachází rostrálně od MI a je podobně organizována. Podrobným mapováním zde také bylo identifikováno somatotopické uspořádání. Největším rozdílem je komplexita pohybů, které lze její stimulací vyvolat. Tam, kde lze stimulací MI vyvolat „pouze” opozici palce a ukazováku, dojde při stimulaci premotorické oblasti např. ke komplexnímu pohybu, který napodobuje psaní. Komplexnost pohybů se zvyšuje s tím, čím rostrálněji probíhá elektrická stimulace. Tyto experimentální nálezy vedly k teorii, že v rostrální části premotorické oblasti vzniká tzv. motorický obraz.
Motorický obraz je v podstatě vnitřní reprezentace pohybu, jakási vizualizace toho, které svaly a s jakou intenzitou musí být aktivovány. Poté je motorický obraz v kódované formě veden asociačními vlákny kaudálně a s každým přepojením probíhá rozklad původního obrazu na dílčí pohyby (přesněji jejich přesně načasovanou sekvenci), které musí být uskutečněny k dosažení požadovaného výsledku. Jakmile dosáhne motorický obraz kaudálního konce premotorické oblasti, je přepojen asociačními vlákny do primární motorické oblasti (MI), jež odpovídá za exekuci pohybu – aktivuje příslušné míšní neuronální okruhy (vzácně pak přímo alfamotoneurony, viz výše).
Premotorická oblast současně vysílá descendentní projekční vlákna do podkorových oblastí, zejména pak do bazálních ganglií, jež přes svůj okruh a talamus zpětně regulují aktivitu v celé motorické kůře.
V premotorické oblasti se také nacházejí tzv. zrcadlové neurony, funkčně velmi zajímavé, neboť se aktivují jak při provádění specifických pohybů, tak i při pozorování jiných jedinců, kteří pohyb konají. Má se za to, že zrcadlové neurony nám umožňují pochopit chování dalších jedinců a učit se pozorováním.
Suplementární motorická oblast
Suplementární motorická oblast leží zejména na mediálním povrchu hemisféry skrytém ve fissura longitudinalis (s. fissura intrahemispherica). Stimulace této oblasti vede k bilaterálním chápavým pohybům rukama, které jsou nejspíše evolučním rudimentem nutným pro pohyb ve větvích. Existuje obecná domněnka, že se do jisté míry účastní tvorby motorického obrazu. Dosavadní experimenty poukázaly na její význam při plánování fixačních pohybů.
Převod signálu z motorické kůry ke svalům
Převod signálu z motorické kůry ke svalům nastává dvojím způsobem: buď přímým převodem kortikospinální dráhou až k alfamotoneuronům, nebo nepřímou cestou přes mozeček, bazální ganglia a různá jádra kmene.
Pyramidová dráha
Pyramidová neboli kortikospinální dráha začíná na:
1) Velkých neuronech motorické oblasti (MI), tzv. Beckových buňkách – asi 30 % vláken
2) Pyramidových neuronech premotorické a suplementární oblasti – asi 30 % vláken
3) Pyramidových neuronech somatosenzorické oblasti (SI) – asi 40 % vláken
Procházejí mezi putamen a nc. caudatus v tzv. capsula interna. Poté se tříští do fibrae pontis longitudinales (též roztříštěné svazky pyramid) a spojují se až u decussatio pyramidum, kde se i většina vláken kříží. Tato zkřížená vlákna tvoří laterální kortikospinální trakt, který postupuje dolů a končí na neuronech příslušného segmentu (alfamotoneuronech, ale i interneuronech). Vlákna, jež se v decussatio pyramidum nezkřížila, formují tzv. ventrální kortikospinální trakt. Tato vlákna se obvykle zkříží později v některém hrudním segmentu. Přispívají zejména k posturální motorice.
V celém svém průběhu vysílá pyramidová dráha četné kolaterály do nejrůznějších struktur včetně bazálních ganglií, mozečku či nucleus ruber.
Rubrospinální trakt
Nucleus ruber, které se nachází v mesencefalu, nabízí alternativní dráhu pro převod signálu do neuronálních poolů v míše. Funguje v blízké asociaci s kortikospinální dráhou.
Nucleus ruber získává informace dvěma cestami:
1) Přímo z kůry – tzv. tr. corticorubralis
2) Kolaterálami z tr. corticospinalis
Tato vlákna končí v tzv. magnocelulární části nc. ruber, kde se nacházejí neurony jen o málo menší než Beckovy pyramidy. Jejich axony pak dávají vznik rubrospinální dráze. Vlákna tr. rubrospinalis končí převážně na interneuronech tvořících součásti příslušných neuronálních míšních okruhů pro daný pohyb.
Zajímavé je, že magnocelulární část nc. ruber obsahuje somatosenzorickou reprezentaci všech svalů (motorického homunkula) podobně, jako motorická kůra. Existuje úvaha, že kortikorubrospinální systém slouží jako záložní okruh, který se uplatní zejména při poškození pyramidové dráhy. Tuto funkci opravdu dokáže vykonávat, jedinou výjimku tvoří jemné pohyby prstů, které jsou při lézi tr. corticospinalis ztraceny.
Vzhledem k tomu, že jak pyramidová, tak rubrospinální dráha probíhají laterálními provazci míšními a inervují zejména míšní okruhy nebo alfamotoneurony pro distální svalstvo, jsou často souhrnně nazývány laterálním systémem motoriky.
Kromě korových oblastí existují i další dráhy známé jako extrapyramidové dráhy. Od tohoto pojmu se dnes ustupuje kvůli jeho malé přesnosti. Bývaly sem řazeny tr. reticulospinalis, tr. vestibulospinalis, tr. tectospinalis a některými autory dokonce i tr. rubrospinalis. Společné měly to, že se nacházejí pod větší kontrolou mozečku a bazálních ganglií.
Mozeček
Cerebellum neboli mozeček je oblast, která zodpovídá za tak důležité funkce, jako je časování a kontrola průběhu pohybu, stejně jako za motorickou paměť. Tyto funkce mozečku byly objeveny teprve nedávno, a proto býval nazýván „tichou oblastí” mozku, neboť jeho elektrická stimulace nikdy nevedla k žádnému výsledku, a to z toho důvodu, že přímo nezasahuje do inervace svalů.
Mozeček získává v každém okamžiku informace o průběhu pohybu jak z vyšších motorických center, tak z periferních receptorů. Tyto informace mezi sebou vzájemně srovnává a při neshodě provede přenastavení těch částí motorických center, které odpovídaly za plánování pohybu. Jinými slovy: mozeček v každém okamžiku porovnává skutečný průběh pohybu s motorickým obrazem. Pokud si neodpovídají, provede korekci center, ve kterých motorický obraz vznikl. To vše na podvědomé úrovni. Dále sám přispívá do plánu pohybu tím, že kůře pomáhá s časováním jednotlivých částí motorického obrazu. Provádí to tak, že propočítává průběh dalšího pohybu v sekvenci ještě během exekuce předchozího. Navíc jeho časovací funkce má autokorektivní funkci. Ta je součástí motorického učení a spočívá ve změnách synaptické plasticity dle toho, zda se propočty časování rovnaly vlastní exekuci pohybu. Jinými slovy, mozeček se sám učí časovat pohyby vlastními chybami. Čím častěji pohyb provádíme, tím lepší a plynulejší je jeho exekuce.
Funkční jednotka mozečku
Mozeček je složen z asi 30 miliónů funkčních jednotek, jejichž základem je pár Purkyňova buňka – buňka hlubokého jádra.
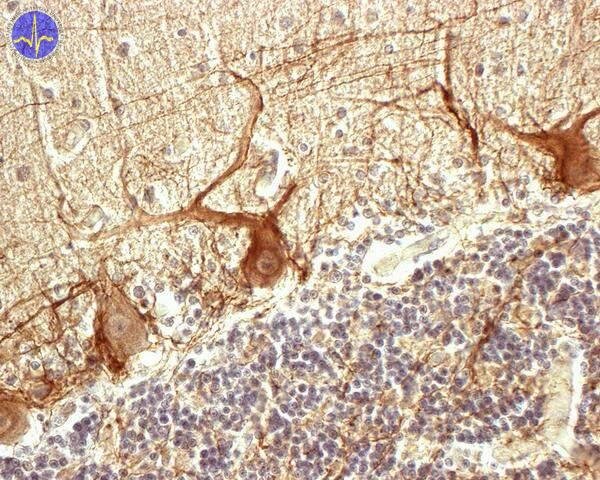
Protilátka značí neurofilamenta v Purkyňových buňkách a axonech košíčkových buněk
Do mozečku přichází dvojí aference – šplhavá vlákna a mechová vlákna.
Šplhavá vlákna
Všechna šplhavá vlákna přicházejí z nc. olivaris inferior. Každé šplhavé vlákno sdílí synapse jak s buňkou jádra mozečku, tak s dalšími 5 – 10 Purkyňovými buňkami (na jedné z nich vytvoří až 300 synapsí). Šplhavá vlákna tvoří zvláštní typ vzruchu zvaný komplexní spike. Lze jej naměřit na Purkyňových buňkách a začíná jedním výrazným hrotem, jejž následuje dlouhá série postupně slábnoucích hrotů. Purkyňovy buňky jsou obrovské inhibiční neurony, které silně snižují aktivitu buněk hlubokých jader.
Mechová vlákna
Mechová vlákna přicházející do mozečku z kůry, kmene a míchy excitují jednak hluboké neurony mozečkových jader, jednak stovky granulárních buněk ve stratum granulare kůry mozečku. Tyto granulární buňky jsou excitační interneurony, které vysílají paralelní vlákna do stratum moleculare mozečku. Paralelní vlákna (nalezneme jich zde miliony) pak tvoří synapse s dendritickým stromem Purkyňových buněk. Aby byla Purkyňova buňka excitována, musí být zároveň aktivních mnoho paralelních vláken (všimněte si rozdílu oproti jedinému mechovému vláknu). Zajímavé je, že aktivita v paralelních vláknech vyvolá na Purkyňových buňkách jinou odezvu, která se nazývá jednoduchý spike. Ten je mnohem kratší než spike komplexní, a nevykazuje proto takovou inhibiční aktivitu na buňku hlubokého jádra.
Takto zapojená funkční jednotka mozečku vede ke křehké rovnováze mezi excitační a inhibiční aktivitou na buňky hlubokých jader, z nichž pak je vedena zpracovaná informace ven z mozečku.
Jak mechová, tak šplhavá vlákna mají kolaterálu, která přímo stimuluje neuron hlubokého jádra. Zároveň oba typy vláken stimulují inhibiční Purkyňovu buňku.
Funkce mechových vláken
Při běžné motorické činnosti převládá aktivita v mechových vláknech. Ta vede ke stálé lehké převaze excitačního působení na buňky hlubokých jader a aktivita jader zůstává konstantní.
Při rychlé sekvenci pohybů přichází informace o motorickém obrazu z kůry prostřednictvím mechových vláken. Při každém jednotlivém pohybu, který tvoří motorický obraz, je nabuzena příslušná část hlubokých jader, a to přímou kolaterálou z mechových vláken. Následuje aktivace mnoha paralelních vláken, jež excitují Purkyňovy buňky a ty aktivitu hlubokého jádra ukončí. Takto probíhá časování pohybu na úrovni funkční jednotky mozečku. Následný inhibiční signál se nazývá „damping”.
Funkce šplhavých vláken
To, jak rychle bude docházet k časování pohybů, musí být mozečkem naučeno. Jak již bylo řečeno, mozeček srovnává vstupy z motorického obrazu s opravdovým průběhem. Zde vstupují do hry šplhavá vlákna. Za normálních okolností vysílá oliva inferior jen asi vzruch za sekundu. Pokud je vykonávaný pohyb prováděn nově, dochází ke zvyšování této frekvence, neboť se zvyšuje i pravděpodobnost chyb. Šplhavá vlákna tak vysílají určitý „error signál”. Komplexní spike se šíří v Purkyňově buňce oběma směry a obnovuje iontové složení i v nejvzdálenějších částech dendritického stromu (tzv. efekt smazání tabule). Ten značně usnadňuje průběh změn ve struktuře některých synapsí. Přesný mechanismus ale není znám.
Bazální ganglia
Bazální ganglia jsou dalším přídatným motorickým systémem. Existují dva významné okruhy, které zprostředkují různé funkce, a to:
1) Okruh přes putamen
2) Okruh přes caudatum
Okruh přes putamen
Jednou z hlavních rolí bazálních ganglií je kontrola komplexních naučených motivů v motorickém obrazu. Jinak řečeno, kontrolují, zda jsou jednotlivé pohyby naplánovány s náležitou intenzitou a ve správném pořadí, zejména u pohybů, jež bylo potřeba se naučit. Jako příklad se často uvádí psaní. Jedná se o velmi komplexní pohyb striktně kontrolovaný bazálními ganglii. V případě poškození ganglií je psaní velmi obtížné, pomalé a postrádá základní úpravu. Výsledné písmo vypadá, jako kdybychom psali poprvé. Právě tuto úlohu zprostředkovává okruh přes putamen.
Okruh přes putamen začíná v premotorické, suplementární motorické a somatosenzorické kůře. Všimněte si, že nevychází z primárního motorického komplexu. Vlákna z těchto oblastí vstupují do putamen, z něj do globus pallidus, následně do VA a VL jader talamu a konečně prostřednictvím talamokortikální projekce zpět do výchozích korových oblastí, a dokonce i do primárního motorického komplexu.
S tímto okruhem funkčně spolupracuje tzv. doplňkový okruh bazálních ganglií, v podstatě pouhá odbočka okruhu přes putamen. Vede z putamen do globus pallidus externus a následně přes subtalamus a substantia nigra do motorického kortexu.
Okruh přes caudatum
Většina naší motorické aktivity pochází z vědomého rozhodnutí. Proces, ve kterém se toto rozhodnutí utváří, se nazývá kognitivní kontrola motorické aktivity. Z velké části za ni odpovídá právě okruh přes caudatum.
Do kaudáta vstupují projekce z většiny částí kůry. Z kaudáta pak vycházejí vlákna do globus pallidus, následně do VA a VL jader talamu a z nich do prefrontální, premotorické a suplementární motorické oblasti.
Pokud například máme hlad, je tato informace zpracována spolu s dalšími senzorickými vjemy v okruhu bazálních ganglií, což nakonec může vyústit v to, že se zvedneme z gauče, zamíříme k lednici a něco si vezmeme (vlastní motorická činnost při obstarání potravy).
Autor podkapitoly: Patrik Maďa
![]()