Content:
1. Introduction to motor functions
2. Reflexes and muscle receptors
3. Central control of motor functions
_
Introduction to motor functions
Our central nervous system gathers and assess sensoric informations. In cooperation with this it has to be equipped by systems enabling to react properly to those informations. The most common reaction to the majority of stimuli is motor response.
This trend is common to all animals across taxons. Because of this hydra contracts after mechanical irritation, deer escapes after even merest trace of danger or some hungry predator goes out to look after a prey.
_
Reflexes and muscle receptors
Spinal cord reflexes
In this part we will engage in spinal cord reflexes. Reflex is the basic unit of the motor activity. Morphologic base of reflexes are specialised neuronal circuits, which are coordinating function of muscles to create effective movements. Without these circuits are even the most simple movements impossible.
The role of higher motor centres are based mostly on activation of spinal cord reflex circuits with according intensity and in correct sequence after stimuli arrive to centres. After the spinal cord cut off we are unable to walk even when higher centers functions are remaining because motor areas of the cortex has no access to activation of neural spinal cord circuit involved in walking.
Reflex arc
Basic arrangement of the spinal cord reflex is called reflex arc:
1) Sensor
2) Afferent pathway (centripetal)
3) Centre of reflex
4) Efferentní pathway (centrifugal)
5) Effector
The basic principle of the reflex arc is simple. Sensor takes adequate stimulus and sends information to the centre of this reflex where is this information evaluated and integrated with other adequate stimuli. Effector finally mediates according motor response.
Sensor
Every receptor apparatus which reacts by response to irritation is considered to be a sensor. Free neuronal endings registering pain or Golgi bodies in muscle tendon are two examples from large group of sensors.
Afferent pathway
Afferent pathways are build up by neural fibers, majority of them comes from the neural cells of spinal ganglia. They trail from the appropriate receptor and enter the CNS by dorsal horn of spinal cord. They spreads in the spinal cord to two collaterals:
1) Collateral ending on the body and dendrites of gray matter neurons of according spinal segment – induces spinal reflexes
2) Collateral ascending to the higher level of the CNS and ends in more distant spinal segments or on neurons of the brain stem or of cortex (usually in sensoric areas)
Center of reflex
In this part we will be talking about simple reflexes, so now (but just now) we can say that reflex center is located in the gray matter of the spinal cord. Here are few millions of neurons which accept informations from sensoric systems. Motor neurons of frontal horn and interneurons are very important for motor reflexes.
1) Motor neurons
They are big neurons and their axons are called efferent fibres in reflexological terminology. We can divide them to groups again, alpha-motoneurons and gamma-motoneurons are the most important.
a) Alpha-motorneurons
Alpha-motoneurons cause activation of effector. They have bulky myelinated axons conducting signals very fastly. They are branching in their ending of axon and finishes by neuromuscular junction at muscle fibres. One motor neuron innervates many muscle fibres usually, these contract together when is stimulated. Alpha-motoneurons are directly responsible for the activation of an effector.
All of the muscle fibres innervated by the same alpha-motoneuron constitute motor unit. Motoric unit is very important term in physiology. Because the power of a contraction is dependent just on the number of activated motor units. The number indicates the sensitivity of motor control of a concrete muscle. For example m. quadriceps femoris adapted to coarse movements is composed of big motor units. On the contrary oculomotor muscles motor units are small and alpha-motoneurons usually innervates single fibres and so allow to manage accurate control of movement to fractions of millimetre.
b) Gamma motor neurons
These motor neurons are significantly smaller than alpha-motoneurons and their axons ends on special muscle fibres called intrafusal fibres which create peculiar muscle spindle participating on the control of muscle tone. For e detail see below.
2) Interneurons
Interneurons are situated diffused in the whole spinal cord segment and their count is thirty time greater than of motor neurons. They are plentifully connected between and create a complicated network which serves for integration functions of the spinal cord. This network of related neurons we call neuronal pool and the motif in which they are connected we call neuronal circuit. Circuits can have these characteristics: they are diverging, converging, reverbing, with serial processing or with parallel processing.
a) Divergence
Divergence is understanded as spreading of signal from one neuron to a whole range of subsequent neurons. So information is transmitted from a one neuron to at least two further sharing synapses with the first one.
b) Convergence
Convergence is situation when a one neuron accepts information from at least two another neurons simultaneously.
c) Reverberation
Reverberation is situation when one information circles in a neuronal circuit repeatedly. It can be reached by neuronal collateral linked to previous neural cell or by many of neurons with serial processing building own closed circuit (repetitive circuit).
d) Serial processing of information
This is situation in which a one neuron transmits appropriate information to just another one neuron and so on. Neurons are creating continuous chain.
e) Parallel processing of information
A parallel transmission means when the same information is managed by at least two neurons in a one moment and whenever is a consequence of a preexisting divergence. Thanks to this a one stimulus can activate more effectors. For example a tread on some sharp object (pain stimulus) activates flexion of injured limb and extension of contralateral.
The most of incomes from higher levels of the CNS are ended at appropriate neuronal pool. In the same exist a small amount of reflex circuits ending on a motoneuron. They are very important so we will discuss the below.
Efferent track
Efferent track goes out from the anterior horn of the spinal cord and consists of motor neurons axons ending on an appropriate effector.
Effector
Is an organ constituted by excitable cells and its function regulates corresponding reflex circuit. Within the purview of the motor system consists of a striated muscle but is very important to realize that exist visceral reflexes with effectors as smooth muscle or various glands.
Muscle receptors
Before we will continue in describing of basic principles of motoric control we have to mention that for correct function of the motion apparatus is feedback monitoring of actual muscle situation as important as activation of motor units. Informations as a change in the length or tension in an exact muscle are gathered by two highly specialised sensory receptors:
1) Muscle spindle
2) Golgi tendon organ
Muscle spindle
Muscle spindles consists of modified muscle fibres, so called intrafusal fibres, which maintain contractile attribute but parallelly serve as receptors of a muscle length and change of length. They are tiny and the whole muscle spindle comprises 4 to 12 intrafusal fibres. Muscle spindles are connected parallely to own muscle by glycocalyx of myocytes. Normal fibres of this muscle are called extrafusal fibres.
Intrafusal fibres differs from extrafusal by central part which doesn’t have so much contractile elements and serves just as area for reception. Stimulus is achieved by stretch of intrafusal fibers central portion. Own contraction of this fibre originates just in the border parts of the fibre and this part is innervated by gamma motor neuron.
According to the arrangement is muscle spindle activated in two situations:
1) Stretching of extrafusal fibres to which is attached – so receptor parts of muscle spindles fibres are stretched too
2) Increasing activity of gamma motor neuron which contracts border parts of the muscle spindle fibre and receptor portion is so stretched too again
These changes are recorded by dual constitution of sensoric neural endings:
1) Primary endings (annulospiral fibres)
2) Secondary endings
As appellation indicates annulospiral neural endings twine around intrafusal fibre and we can find theme in the middle of the fibres length. Secondary neural endings have shrubby structure and locate on the borders of the spindle.
Dual arrangement evinces oneself intrafusal fibres. We distinguish:
1) Fibres with nuclear chain
2) Fibres with nuclear bag
These differ by organization of cell nuclei in the central receptor part. Fibres with nuclear chain have their nuclei in a row and create fibre with not so changing diameter than fibres with nuclear bag where nuclei are accumulated in the central part and so make marked bulg.
This morphological phenomenon is very important for function. Fibre with nuclear chain stretching will cause narrowing in the whole length and so activates both primary and secondary neural endings. In the contrary fiber with nuclear bag have enough plasmatic membrane excess in the central portion so there is not such narrowing at the ends of the intrafusal fibre. So there will be change of the diameter just in the neighbourhood of the nuclear bag (where are situated annulospiral endings) but not at borders scanned by secondary neural endings. Fibres with nuclear bag don’t activate secondary neural endings and allow two types of response – static and dynamic.
Static response
Static response is induced when primary and secondary neural endings are activated simultaneously. This phenomenon is associated with slow stretch of a muscle. It informs in every moment about actual stretch of a muscle. The extent of stretch is in direct proportion to the frequency of discharges in both primary and secondary neural endings.
According to how much is intrafusal fibre stretched in the moment is higher frequency of produced action potentials. Centrum of reflex (and higher motor centres) are obtaining correct coded information.
Dynamic response
Dynamic response is reaction to a quick stretch of a muscle. Result is excessive activation of primary nerve endings because of abrupt change of the nuclear bag diameter. This reaction of a muscle spindle associated with predominant activity in annulospiral neural endings is call dynamic response and monitors the swiftness of the stretch change. After the end of the contraction starts simultaneous activity in primary and secondary fibres to get informations about actual length of the muscle again.
The dynamic response is characterised by much higher frequency of action potentials at primary endings than static response.
Regulation of response intensity
Higher neural centers – especially bulboreticular area (which is under the influence of the cerebellum, basal ganglia and motor cortex) – affect sensitivity of the muscle spindle by the change of gamma motor neurons activity. The change in the frequency of discharges leads to contraction of intrafusal fibres and so to their stretch. This increases sensitivity of the central receptive field.
Bulboreticular area is even able to change the sensitivity of dynamic and static response separately. Two types of gamma motor neurons are found: gamma-d mostly innervating fibres with nuclear bag and so modulating dynamic response and gamma-s founded on fibres of nuclear chain involved in the static response.
Muscle stretch reflex
Anytime is muscle stretched rapidly comes reflex contraction. This phenomenon is called the muscle stretch reflex and it is the most simple manifestation of the muscle spindle function. It is monosynaptic reflex dependent just on the dynamic response.
Rapid stretch of muscle evokes high frequency activity in annulospiral fibres. They enter the appropriate segments dorsal horn of the spinal cord then continue and end in ipsilateral anterior horn on the alpha motor neuron of the proper muscle. Following rapid activation leads to equal contraction (“tug”). Role of this reflex is to compensate rapid and unexpectable changes of the stretch.
Imagine situation when you carry a box with books and somebody suddenly throw in some books. In this moment is especially m. biceps brachii rapidly stretched but just in a fraction of second reaction of muscle stretch reflex is this stretch compensated. So you will maintain this holding of the box which didn’t fall.
The reason for which the box is kept in hands even after the extinction of the excessive dynamic response is because of higher activity also in the static response after the rapid compensation. This holds a higher activity in alpha motor neurons of m. biceps brachii. Thanks to this some authors state that muscle stretch reflex has two components: dynamic stretch reflex and static stretch reflex.
So the static stretch reflex replaces the dynamic stretch reflex with a small delay which is based on the whole character of the response (lower frequency, activity also in secondary neural endings) and because of the fact, that on alpha motor neurons end just axons of primary endings. Secondary endings end just on some interneurons and are processed by appropriate neural circuits.
Clinical correlation:
Muscle stretch reflex is used in clinical praxis in neurology. It is simple and reliable test of the reflex sensitivity and so of the CNS correct function.
Role of the muscle spindles in voluntary motor function coordination
Anytime when the execution of some movement is started are simultaneously activated gamma motor neurons (so called coactivation alpha-gamma). Thanks to this are equally contracted both extrafusal and intrafusal fibres. A muscle spindle is proportionally stretched in every moment to a monitored muscle and can fulfil its receptor function.
Muscle spindles are involved in the stabilisation of the position during the motor activity. We can find out higher activity in bulboreticular area linked to any motor operation. This is not common alpha-gamma coactivation. There is automatic sensitisation of spindles in both antagonist and agonist of this movement. Thanks to this are shortened spindles of opposite muscles leading to significant stabilisation of the appropriate joint. Senzitisated muscle spindles can elicit the muscle stretch reflex even after just a little deviation of the position and so can compensate it.
Golgi tendon reflex
This is very important monosynaptic reflex. The sensor is Golgi tendon body incorporated into muscle tendon fibres of the muscle attachment. Thanks to this serial placement with monitored muscle can’t get information of the muscle length. It measures muscle tension. Golgi tendon body informs about both the dynamic and the static response alike the muscle spindle, but the principle is less complicated. Changes in muscle stretch are coded as high frequency activity and steady state is expressed as lower frequency impulses.
The afferent fibre of this reflex circuit enters the CNS by the dorsal horn of the appropriate spinal segment and ends on the alpha motor neuron of the proper muscle. In the contrary to the muscle spindle reflex it is inhibitory synapse. It’s important to emphasize that both afferents of the Golgi tendon reflex and muscle stretch reflex are sending collaterals that end in higher levels of the CNS so motor centres have actual informations about status of every striated muscle in every moment..
This reflex function is in the protecting of the musculature which otherwise can be destroyed by too big tension. When the critical threshold for the Golgi reflex activation is overcomed the appropriate alpha motor neuron is inhibited just in fragment of second and this leads to the relaxation of this muscle, this is called lengthening reaction. Another very important function of the Golgi reflex is to distribute tension evenly to all involved fibres. There is a group of Golgi bodies in every tendon perceiving the distribution of tension in the proper muscle. So if some muscle fibres are overloaded Golgi reflex inhibits just alpha motor neurons of appropriate motor unit and this tension is redistributed within this muscle.
Flexor reflex
Flexor reflex is an example of the polysynaptic reflex. It is also called nociceptive reflex because sensors are nociceptors and the most important trigger is pain. Painful stimulus is conducted by afferents to the dorsal horn of the spinal cord where ends on present interneurons. This activates the whole appropriate neural circuit which sends informations to higher levels of the CNS and elicits local reactions in involved spinal cord segment. This activation of many interneurons leads to:
1) Activation of alpha motor neuron of the proper flexor
2) Inhibition of alpha motor neuron of the opposite extensor (antagonist)
3) Propagation to surrounding spinal cord segments
4) Activation of the crossed extensor reflex
Simultaneous inhibition of an antagonist defends against the eliciting of the stretch reflex which will be provoked otherwise after rapid involuntary activation of flexor. There is finally escape from the painful stimulus and thanks to this is reflex characterised as defensive.
We can state a contact with a hot object as an example. If you are preparing dinner and touch heated frying pan, there is involuntary reaction, which retreats zour upper limb from this painful stimulus. If the antagonist will not be inhibited simultaneously, muscle will be rapidly stretched and so muscle spindle would be stretched too. Stretch reflex alone would compensate the change and press your hand back to the pan.
Crossed extensor reflex
Crossed extensor reflex is another one polysynaptic defensive reflex and is connected to the flexor reflex. It continues automatically 0,5 s after the flexor reflex. This delay is caused by multiple connections in a neuronal pool of an appropriate spinal cord segment. It’s function is the extension of the contralateral limb (contrary to the affected limb so called ipsilateral) In humans it is very important especially for the injury of lower limbs, but it can be provoked at upper limb too. Extension of contralateral limb prevents falling, which should continue if there would be just flexion of the ipsilateral limb.
Now we can imagine, that when the foot is stepping on some nail, at first there is provoked flexor reflex which brings back by rapid activation of the flexor group foot from the harming object. Then follows extension of the contralateral limb and a weight is transferred on it. If there wouldn’t be this extension, both limbs will be in flexion in the same moment and fall will continue. The most probable in this moment is fall on the mentioned nail.
In complete, upper limbs are involved in this reaction too, because this stimulus is propagated to more distant segments, as mentioned before. There are elicited both flexor and crossed extensor reflexes, but side inversed to lower extremities. This means that if there would be provoked flexor reflex for example at lower right limb, there will be extension of upper right limb and left upper limb will be involuntary flected in the contrary. This synchronic activation of couple flexor – crossed extensor reflex of upper extremities is more weak than of lower limbs, where sensors of this reflex were activated.
This phenomenon contributes to better distribution of weight and so to keep balance. In ontogenesis it is probably remnant of quadruped state in the development of our extraordinary organism – human been.
_
Central control of motor functions
Until now, we have discussed at the most about various reflexes which are involuntary and it is impossible to influence them by our will, as it is written in their substance of word. We will describe voluntary motor functions in following text.
Motor cortex
Majority of motor functions is initiated by cerebral cortex. This part of the CNS is sending a planned sequence of directions to neural circuits of the spinal cord, which are activated in appropriate succession. But there are exceptions, when the motor cortex activates straight alpha motor neuron of some muscles, muscles with small motor units are involved in this usually, those needs precise voluntary control as we can see on hand muscles.
The motor cortex is located in the front of the sulcus centralis and occupies about one third of the frontal lobe area. We can divide this motor cortex to three areas with sundry functions:
1) Primary motor area
2) Premotor area
3) Supplementary motor area
Primary motor area
Primary motor area (MI) – area 4 in Brodmann’s classification – is located at the gyrus praecentralis and extends laterally to the fissura Sylvii. Patients undergoing neurosurgical operation were stimulated in this part of brain in the past. It was discovered, that MI is strictly somatotopically organized and its sections we can assign to body parts. This local distribution we call as motor homunculus, because we can project whole human body as a figure to the g. praecentralis to represent location of appropriate motor control center for a part of the body. This figure is considerably deformed, because districts of the cortex are proportional to a level of innervation and not to real size of parts (homunculus biggest districts are hands). And similarly was discovered, that pyramidal neuron does not command one muscle, but controls one specific movement. E.g. opposition of a thumb and an index finger.
Premotor area
Premotor area is located rostrally to the MI and its organisation is similar. It was identified somatotopic arrangement by detailed mapping. The biggest difference is complexity of movements which can be elicited. In the site where we can provoke “just” opposition of a thumb and an index finger at the MI, equal part at the premotor area produce an complex movement e.g. imitating writing. The complexity of movements is more higher according to how much rostrally is electrically stimulated. Those experimental findings led to the theory, that in rostral part of premotor area origins so called motor image.
Motor image is internal representation of movement in substance, some visualisation of which muscles and with how big intensity have to be activated. Afterwards is motor image in coded form transmitted by associational fibres caudally and with every switch is image dispersing to particular movements (more accurately – precisely timed sequence), which has to be realized to achieve demanded result. After arrive to the caudal end of the premotor area is signal transmitted by associational fibres to the primary motor area (MI), which is responsible for the execution of the appropriate movement – activates corresponding spinal cord circuits (in rare cases alpha motor neurons, see before).
Premotor area is simultaneously sending descendent projectory fibres to subcortical areas, especially to the basal ganglia which are by its own circuit and the thalamus retrogradely transmitting activity of the whole motor cortex.
In premotor area are located so called mirror neurons. These function is very interesting, because they are activated by both executing of specific movements and observation of another individuals, which are doing this movement. We consider them as responsible for understanding of behavior of other individuals and for learning by observation.
Supplementary motor area
SUpplementary motor area lies mostly at medial surface of hemisphere covered in the fissura longitudinalis (seu interhemispheric fissure). Stimulation of this area leads to bilateral taking movements of hands which are probably rudiments of motion at tree branches. We thought, that they are involved in creating of the motor image. Experiments to nowadays have shown the importance in the planning of fixation movements.
Transmission of signal from motor cortex to muscles
Transmission of a signal from the motor cortex to muscles is carried out by two different ways. By direct transfer by corticospinal tract to alpha motor neurons or by indirect way across the cerebellum, basal ganglia and various nuclei of the brain stem.
Pyramidal tract
Pyramidal aka corticospinal tract start on:
1) Big neurons of the motor area (MI), which are called Becks cells – counts about 30 % of fibres
2) Pyramidal neurons of the premotor area and supplementary area – counts about 30 % of fibres
3) Pyramidal neurons of the somatosensory area (SI) – counts about 40 % of fibres
They go between the putamen and nc. caudatus in so called capsula interna. Then they are spreading to the fibrae pontis longitudinales (dispersed bundles of pyramids) and then form at decussatio pyramidum, where the most of these fibres cross. Continuing crossed fibres form lateral corticospinal tract, which continues downward and ends on neurons of the appropriate segment (on alpha motor neurons and on interneurons too). The minority of fibres which are not crossed at decussatio pyramidum forms so called ventral corticospinal tract. Those fibres usually cross afterwards in some segment of thorax. They support mainly the postural motor functions.
In the whole course sends this pyramidal tract various collaterals to different structures including basal ganglia, the cerebellum or the nucleus ruber.
Rubrospinal tract
The nucleus ruber, located in the mesencephalon, offers alternative pathway to this transmission of signal to neural pools of the spinal cord. Its function is closely associated to the corticospinal pathway.
Nucleus ruber gains informations by two pathways:
1) From the cortex – so called corticorubral tract
2) By collaterals from corticospinal tract
This fibres ends on so called magnocellular part of the nucleus ruber where we can find neurons just little bit smaller than Becks pyramids. Their axons afterwards give rise to the rubrospinal tract. Fibres of this rubrospinal tract end mostly on interneurons which are parts of appropriate neural spinal cord segments for the corresponding move.
Interesting is, that magnocellular part of the nucleus ruber contains somatosensory presentation of all muscles (motor homunculus) similar to motor cortex. We think that corticorubral system is serving as a reserve system which we apply especially when the pyramidal tract is damaged. It is really able to carry out this function with only one exception of soft finger moves, which are lost in the state with lesion of tractus corticospinalis.
According to the fact that both pyramidal and rubrospinal tracts are going through lateral strands of the spinal cord and innervates especially spinal circuits or alpha motor neurons of the distal musculature, they are summary called: lateral system of motor function.
Except the cortex areas there exist another pathways called extrapyramidal system. We recede from using of this term because of less accuracy. Parts counted toward there are: reticulospinal tract, vestibulospinal tract, tectospinal tract and some authors accounts there rubrospinal tract too. Common characteristic of them is that they are under greater influence of the cerebellum and basal ganglia.
Cerebellum
The Cerebellum, so called little brain, is part responsible for so important functions as timing, control of movement development and motor memory. These functions of the cerebellum were found recently, and so before this discovery was called “silent part” of the brain, because its electric activity never led to any outcome. That is thanks to the fact, that it is not involved in innervation of muscles directly.
The cerebellum in every moment obtains informations about progress of movements from higher motor centres and peripheral receptors. Those informations compares together and when finds mismatch then sets those parts of motor centres, which are responsible for planning of an appropriate movement. By other words, the cerebellum in every moment compares real progress of a movement with motor image. If they do not agree, it makes correction of centres in which the image emerged. Everything mentioned does at the subconscious level. It also contributes to planning of movement by helping cortex to time individual parts of motor image. This performs by calculating of the progress of the next move in the sequence during the execution of the prior move. In addition its timing function has an autocorrect function. This is component of motor learning and is based on changes of synaptic plasticity according to equation of calculated timings to execution of movement. By other words, the cerebellum alone learns the timing of movements by own mistakes. As more frequently we do the movement, its execution is more fluent and better.
Functional unit of cerebellum
The cerebellum is composed of about 30 millions of functional units, unit consists of couple Purkinje cell – cell of deep nucleus.
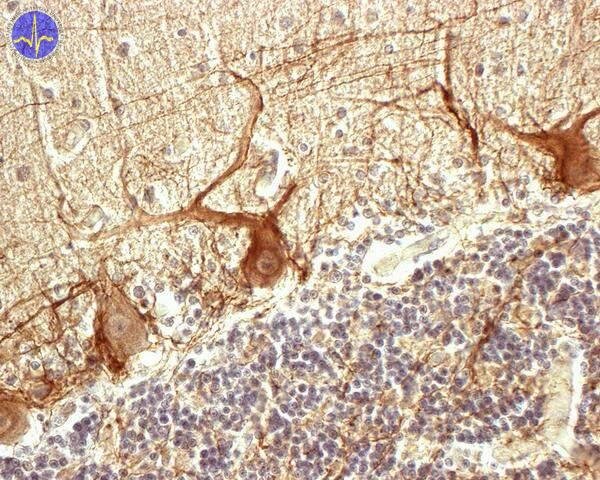
Antibody labels neurofilaments in Purkinje cells and in axons of basket-like cells
To the cerebellum comes two afferents – climbing fibers and mossy fibres.
Climbing fibres
All climbing fibres comes from nucleus olivaris inferior. Every climbing fibre shares synapses with cell of cerebellar nucleus and further 5 – 10 Purkinje cells (fibre makes more than 300 synapses on one). Climbing fibres create a special type of action potential called complex spike. We can measure it on Purkinje cells and it starts by one well marked distinctive spike, which is followed by long series of progressively weakening spikes. Purkinje cells are giant inhibitory neurons, which strongly decrease the activity of deep nuclei cells.
Mossy fibres
Mossy fibres comes to the cerebellum from the cortex, brainstem and spinal cord. This fibres excite deep neurons of cerebellar nuclei and hundreds of granular cells in stratum granulare too. These granular cells are excitatory interneurons, which send parallel fibres to the stratum moleculare of cerebellum. Parallel fibres (we can find there millions of them) then create synapses with dendritic trees of Purkinje cells. Many parallel fibres have to be activated to excite Purkinje cell (notice the difference between just one mossy fibre). Interesting is, that activity in parallel fibres elicite on Purkinje cells another response called simple spike. This is much more shorter than complex spike and so does not evince as big inhibitory activity to the cell of deep nucleus.
This form of functional unit of the cerebellum leads to a fragile equilibrium between excitatory and inhibitory activity to cells of deep nuclei, which transmit this processed information out of the cerebellum.
Both mossy and climbing fibres have collateral directly stimulating neuron of deep nucleus. Both types stimulates inhibitory Purkinje cell simultaneously.
Function of mossy fibres
Common motor activity prevails activity in mossy fibres. It leads to permanent mild predomination of excitatory influence to cells of deep nuclei and activity of nuclei remains constant.
When there is rapid sequence of movements, information about motor image from the cortex comes via mossy fibres. Every single move elicits appropriate part of deep nuclei by direct collateral from mossy fibres. Continue activation of many parallel fibres, which excite Purkinje cells and those end the activity of deep nucleus. This is way how timing takes place on the level of the cerebellar function unit. Following inhibitory signal is called “damping”.
Function of climbing fibres
How fast will be the timing of movements has to be learned. As was mentioned, the cerebellum compares incomes of motor image with real development. Here is important function of climbing fibres. Oliva inferior sends just one excitatory impulse per second ordinarily. This frequency is going to rise when is performing move carrying out first time, because the probability of mistakes rise too. So climbing fibres are sending certain “error signal”. Complex spike spreads through the Purkinje cell to both directions and restores ion composition even in far parts of the dendritic tree (so called effect of table erasure). This considerably ease the course of changes in structure of some synapses. But exact mechanism is not known.
Basal ganglia
Basal ganglia are another accessory motor system. There exist two important circuits arranging various functions:
1) Circuit across putamen
2) Circuit across caudate
Circuit across putamen
One of the most important role of the basal ganglia is the control of complex learned motives of motor image. This means that they control if single moves are planned with appropriate intensity and correct succession. This especially in cases of moves which had to be learned. As an example is usually mentioned writing. It is very complex move which is strictly controlled by basal ganglia. If are ganglia damaged, writing is very difficult, slow and has no basal layout. It looks like affected person writes first time. This task resolves putamen.
The circuit across the putamen starts in the premotor, supplementary motor and somatosensory cortex. Notice that it does not originate in the primary motor complex. Fibres from those areas enter the putamen, continuous to the globus pallidus and then follows to VA and VL thalamic nuclei. and even to the primary motor cortex.
With this circuit cooperates so called accessory circuit of basal ganglia, which is in substance just turning of the circuit across the putamen. It goes from the putamen to the globus pallidus externus and then across the subthalamus and substantia nigra to the motor cortex.
Circuit across caudate
The most of our motor activity is based on our conscious voluntary decision. Process of creating of this decision is called cognitive control of motor activity. The caudate is responsible for the major part of it.
Projections from the majority of cortex enters the caudate. From the caudate then goes fibres to the globus pallidus and continue to VA and VL nuclei of thalamus. From those locations goes to the prefrontal, premotor and supplementary motor area.
If we are hungry, this information is processed with another sensory perceptions to the circuits of basal ganglia, what can lead to getting up from a sofa and going for some food to our fridge (concrete motor activity of getting a nourishment).
Subchapter Author: Patrik Maďa
![]()