Content:
1. Introduction to hearing and vestibular apparatus
2. Hearing
3. Vestibular apparatus
_
Introduction to hearing and vestibular apparatus
In this chapter we will discuss two different senses: hearing and vestibular apparatus. These two are often discussed together because of their anatomical relations.
_
Hearing
Hearing is a sense, which allow us to perceive propagation of a longitudinal waves in the adjacent space. And especially interpret those waves as a sound. It is assumed that this sense evolved as an analyser of surroundings to detect approaching predator. Utilisation of hearing differentiate during evolution. Some groups of animals utilise hearing in a different way than their predecessor did. For example bats use hearing as a main analyser of a surroundings – contrary to humans, whose main analyser is sight. Ears of bats are so sensitive that they can distinguish between fell of male and fell of female. In another species hearing got prime role in registering a communication sounds as is truth for humans and cetaceans.
Acoustic
Acoustic is a branch of science, which study origination and propagation of a sound waves. Presence of molecules is a necessary condition for propagation of a sound. So that we can hear a train approaching, while our ear is enclosed to a rail. Or hear its hoot while standing nearby. But we can’t hear it in a vacuum. Sound is a longitudinal oscillation, which need space with molecules in it to propagate. In fact, sound itself is an alternation between compression and rarefaction of a molecular medium.

Distance between two compressions (or two rarefaction) in a medium is called wavelength. Compressions and refractions spreads through medium with speed v, which is equal to 340 m/s, while the temperature of medium is 0°C. Local changes in density of medium create unstable acoustic pressure. Acoustic pressure acquires high values in compressed medium and low value in rarefied medium. As the waves propagate through medium, acoustic pressure changes periodically. These changes are known as acoustic waves.
Properties of sound can be recorded by sinusoid, where wavelength, amplitude and frequency can be easily described.
Amplitude scales with degree of compression in medium and increment of acoustic pressure. It is subjectively perceive as sound volume. Higher amplitude means higher sound volume.
Wavelength corresponds with strike note. Higher value of wavelength means lower pitch. Because wavelength bear upon frequency, we can postulate that higher frequencies tend to be be lower pitch.
Simple tone is a sound composed of oscillations with same value of frequency. In other words proper simple tone is a sound that can be recorded by only one oscillatory function (sinusoid). These tones aren’t very common in nature. Most of our auditory perceptions are so called overtones. Overtone is a sound composed of many oscillatory functions, which differ in frequencies. Mind you, that these frequencies aren’t random. They possess high degree of organisation. There is one oscillation which has so called fundamental frequency. All the others oscillations – which are composing the sound – acquire values that equal integral multiples of fundamental frequency. These are called harmonic partials. For example diapason A has frequency 440 Hz. Its harmonic partials are 880 Hz, 1320 Hz, 1760 Hz, etc. Tone pitch is identified by the lowest frequency. Harmonic partials identify a timbre. All common sounds around us are composed of overtones, sound of the grass in the wind, voices of people passing by, even your favourite song. And only because of harmonic partials we can tell the difference between diapason A played on a piano and diapason A played on a violin.
Noise is a miscellany of oscillations with different frequencies, most of which are mutually incommensurable. So that oscillations composing noise are completely random. They even differ in amplitudes.
Humans can perceive sounds with frequencies between about 15 Hz and 16 kHz. Sensitivity to higher frequencies decline with age. Upper border can be as low as 5 kHz in elderly people. This phenomena can be seen at train stations. Young children cover their ears while the train is retarding. The elders are not. They don’t feel any discomfort, because they aren‘t able to hear the noise of a breaking train. Human ear is most sensitive to frequencies between about 2 kHz – 4 kHz. These are most important in understanding of speech.
Another important term is an auditory threshold. It is a minimum acoustic pressure of a simple tone that an average ear can hear with no other sound present. Auditory threshold therefore differs for distinct frequencies. If there are another tones aside from tested simple tone, then its auditory threshold significantly increases. This phenomena is called auditory concealment.
Sound volume scales with acoustic pressure. Because of small dimension of acoustic pressure values, new magnitude was found: Sound pressure level (Lp). Its unit is a decibel – dB. And it is defined by following formula:
Lp = 20 log(Px/P0)
Px – immediate acoustic pressure
P0 – auditory threshold – frequency specific
As we can see, if immediate acoustic pressure raise 1000 – fold, then sound pressure level will raise by 60 dB. Because logarithmic expression raised 1000 – fold, we need to know that logarithm of 1000 equals 3. Then Lp = 20 x 3 = 60.
To determine very sound volume, Lp need to be measured for different frequencies and recorded as graph. Recorded frequencies are chosen according to subjective indication of a same sound volume. Graph fond this way is called isophone.
According to isophone we can indicate Lp for each one simple tone. Lp values found this way differ, but are subjectively heard as equally loud.
If isophone connects smallest hearable values of sound pressure level we get isophone for auditory threshold. If the values of Lp are risen high enough during examination, pain threshold is found. At this point forth we do not perceive oscillations as a sound but as a pure pain.
For frequencies about 2 kHz:
4 dB – Acoustic threshold
20 dB – Complete silence
30 dB – Whisper
60 dB – Speech
100 dB – Concert
130 dB – Pain threshold
140 dB – Acoustic trauma
Sound volume is measured during so called subjective audiometry in clinical practise. It is denoted in phons. 1 phon is equal to 1 dB for frequencies between about 2 kHz – 4 kHz.
Sone is another way of measuring sound volume. Number of sones indicates how many fold louder the tone is compared to basic tone. According to definition 1 sone is a sound volume of a tone with Lp equals 40 dB and frequency 1 kHz. 2 sones is value of sound volume twice as loud.
Sound perception
By perception of sound we understand the conduction of mechanical longitudinal oscillation through inner ear and its own registration in the Organ of Corti. Registered information are stored in higher associative centres. These information are subdued to complete analyses and are used to form an inner image of reality.
Outer ear
Main role of outer ear is to capture mechanical oscillations and to transmit them right to the tympanic membrane.
Middle ear
Lateral border of a middle ear is a tympanic membrane. Vibration of a tympanic membrane are transmitted to malleus, then to incus and stapes at last. Base of stapes transmits vibrations to perilymph. It is a system of swipes which prevents a losses of acoustic energy. So that middle ear block its refraction.
Area of tympanic membrane is many fold bigger than basis of stapes. Because a product of pressure and area is constant, the transmission apparatus reduce energy losses. Relatively small acoustic pressure is transmitted to perilymph as a many fold bigger pressure ,which induce a hearing sensation. In other words, middle ear is an amplifier of acoustic pressure. If there was no middle ear ossicles, then majority of acoustic energy would reflect.
There are two muscles anchoring to the middle ear ossicles: stapedius muscle and tensor tympani muscle. Both are contracting during excess sound stimulus to protect sensitive inner ear apparatus. So their main purpose is to act as an attenuator of acoustic pressure.

Inner ear
Inner ear is located in temporal bone cavity called bone labyrinth. Cochlea as well as vestibular system can be found there. Cochlea – a very sound receptor – is a snail like organ with three caverns: scala vestibuli, scala media and scala tympani.
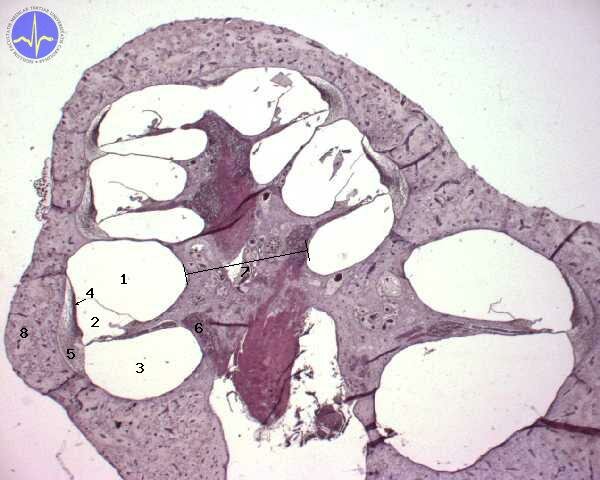
1 – Scala vestibuli, 2 – Scala media, 3 – Scala tympani, 4 – Stria vascularis, 5 – Spiral ligament, 6 – Spiral ganglion, 7 – Modiolus, 8 – Bony cochlea
1) Scala vestibuli adjoints foramen ovale. It contains perilymph. Scala vestibuli is located in on the inner surface of cochlear convolutions.
2) Scala media contains hair cells, which are located on a basilar membrane of organ of Corti. Scala media is filled with endolymph.
3) Scala tympani contains perilymph. Scala tympani can be found on the outer surface of cochlear convolutions. It adjoints foramen rotundum.
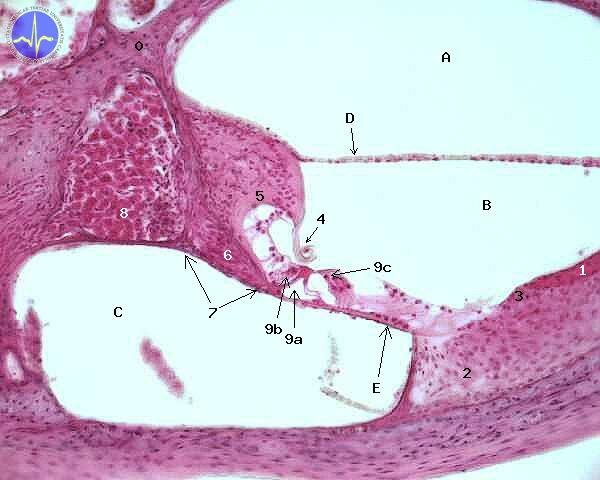
0 – Modiolus, 1 – Stria vascularis, 2 – Spiral ligament, 3 – Spiral prominence, 4 – Tectorial membrane, 5 – Limbus spiralis, 6 – Cochlear nerve fibers, 7 – Osseal spiral lamina, 8 – Spiral ganglion, 9a – Inner tunnel of Corti, 9b – Inner hair cell, 9c – Outer hair cells, A – Scala vestibuli,B – Scala media, C – Scala tympani, D – Vestibular membrane, E – Basilar membrane
Caverns are mutually separated by membranes. There is basilar membrane between scala media and scala tympani, Reissner’s membrane between scala media and scala vestibuli. Tectorial membrane is conjoint with Reissner’s membrane, located in scala media. Tectorial membrane is in a slight contact with hair cells of organ of Corti.
Generation of hair cell action potential
If the basis of stapes start to vibrate, these vibrations are transmitted to perilymph of scala vestibuli. There vibrations spread to Reissner’s membrane and finally to tectorial membrane, which is slightly touching apical surface of hair cells. Mutual movement of tectorial membrane and hair cells is registered by first neuron of auditory pathway.
Apical surface of hair cell is washed by endolymph. Composition of endolymph is similar to the composition of a intracellular fluid. It contains high concentration of potassium (about 150 mmol/l). Concentration high like this is generated by potassium channels of vascular stria. Cells of vascular stria obtain potassium ions by means of Na+/K+/2Cl– symporter. This symporter can be also found in the loop of Henle.
If the hair cell is deflected by the instrumentality of vibrations, the mechanical potassium channels open and cause influx of potassium to the hair cell. Influx of potassium fires off action potential. From hair cell, potassium lekas to the perilymph with low potassium concentration. This process leads to repolarisation. Ions are actively pump over from perilymph to endolymph and the circle is closed.
Clinical correlation:
Increase in permeability of membranes that separate endolymph from perilymph is one of the main mechanisms in pathogenesis of Meniere’s disease. Meniere’s disease is characterised by attacks of hearing impairment.
Genetic defect in potassium channel of vascular stria leads to deafness and long QT interval syndrome. Because this channel is also expressed in myocytes of heart, its defect leads to prolongation of repolarization – which in turn causes long QT interval.
Coding of frequency and intensity of sound
Vibrations spread from foramen ovale to the centre of the cochlea (helicotrema). To differentiate simple tones from each other a properties of basilar membrane are changing during its course. Its rigidity is declining towards the helicotrema. According to physical laws for every oscillation of certain frequency between 15 Hz and 20 kHz there is a part of basilar membrane that will resonate. In other words: for every frequency there is an vibrating maximum – part of basilar membrane with extremely high amplitudes, that are recorded by hair cells. This theory is called phonotopy.
High frequencies have their vibrating maximum near foramen ovale. Near helicotrema lower frequencies are registered. The most vibrating maximums are specialised to registered frequencies between about 2 kHz and 4 kHz. In this interval we can differentiate sounds that differs from each other only by 3 Hz.
Auditory pathway
Auditory pathway consists of four neurons. According to a new research there are many more synapses and its structure is far more complicated.
Because hair cells do not have their own axons, the first neuron is bipolar cell of cochlear spiral ganglion. Its dendrite synapses with hair cell and thus create cochlear part of vestibulocochlear nerve. It enters cochlear nucleuses in the caudal part of pons (pons Varolii). Here it synapses with second neuron and leaves cochlear nucleuses in one of three fascicles:
1) From ventral cochlear nucleus as trapezoid corpus
2) From ventral cochlear nucleus as intermediate acoustic stria
3) From dorsal cochlear nucleus as dorsal acoustic stria
Most of second neurones crosses and form lateral lemniscus. Axons of the lateral lemniscus synapses in inferior colliculus, where the third neuron originates.
There is some discussion if the third neuron isn’t actually the fourth one. Some axones of the lateral lemniscus in fact synapses in superior olivary nucleus, the others in trapezoid corpus nucleus and part of the rest in lateral lemniscal nucleus.
Axones from inferior colliculus enters medial geniculate corpus of thalamus a synapses with the fourth neuron. The fourth neurons form a bundle called acoustic radiation which enters primary auditory cortex located on superior temporal gyrus.
Organisation of the primary sensory cortex as well as organisation of the auditory pathway is tonotopic. Which mean that only certain fibres are activated by certain frequencies. Activated fibres act only in specific locations of primary auditory cortex. So that tones, volume and source of sound can be analysed.
There are association fibres which lead from the primary auditory cortex to a secondary auditory cortex, where the inner representation of a sound is formed. This representation of a sound is then analysed in Wernicke’s area, where the meaning is decoded.
According to some authors it is secondary auditory cortex where auditory memory is stored.
Interestingly unilateral lesions don’t cause serious hearing impairment, because quite big portion of fibres is sent to contralateral pathway at every level of auditory pathway.
_
Vestibular system
In fact vestibular apparatus consists of two distinct sensory organs: static and kinetic sensor. It is located to bone labyrinth as well as cochlea. Static sensor is formed by utricle and saccule. Kinetic sensor is formed by three mutually perpendicular semicircular canals.
All parts of vestibular apparatus are filled with endolymph and their outer surface is washed by perilymph.
Static sensor
There are epithelial prominences called macula in utricle and saccule. On a surface of the macula hair cells are located. Macula is placed perpendicularly to an axis of human body in the utricle. Whilst in saccule, macula is parallel with the axis. Surface of macula is covered in mucopolysaccharidic substance, which contains otoliths. Otoliths are crystalline particles of calcium carbonate (CaCO3) and a protein. If the head is bent backward, forward or sideways the otoliths move and excite hair cells. Therefore static sensor informs about changes of head location with respect to gravitation field. Utricle and saccule hair cells can also be excite by linear acceleration. If the vector of the acceleration lies in vertical plane, the saccule is activated. This happens all the time while taking an elevator. If the vector of the acceleration lies in horizontal plane, the utricle is activated. As it is true for driving a car.
It is important to know, that action potentials are transmitted from utricle and saccule in greater extent during movement of otoliths. If otoliths are still, there is no hair cell excitation and therefore no action potential. Central nervous system uses last known information of the head location, while static sensor is inactive.
Kinetic sensor
Kinetic sensor by three mutually perpendicular semicircular canals: anterior semicircular canal, posterior semicircular canal and lateral semicircular canal. There is an crista ampullaris with hair cells in every canal. Surface of crista ampullaris is covered in mucopolysaccharidic substance with otoliths.
Kinetic sensor register an angular acceleration. During head rotation endolymph is trying to preserve its location according to Newton’s first law. Therefore momentum of endolymph causes it to move with regard to the crista ampullaris, moving the otoliths and exciting hair cells.
Vestibular pathway
Vestibular pathway consist of three neurons. Hair cells of crista ampullaris and makula don’t have their own axons, as is it true for the hair cells of the organ of Corti. Therefore they are innervated by neurons originating in vestibular ganglion. Axons of these neurons form a vestibular part of vestibulocochlear nerve. Some fibres don’t participate in vestibular pathway. They enter cerebellum directly in a direct vestibulocerebellar pathway. So cerebellum disposes of an actual location informations before the consciousness does. The others fibres enter the pons, where vestibular nucleuses are located. They are called: lateral nucleus, superior nucleus, medial nucleus and inferior nucleus. Here the neurones from vestibular ganglion synapse.
Second neurones enter following structures:
a) Cerebellum as indirect vestibulo-cerebellar pathway
b) Spinal cord as vestibulospinal pathway
c) Nucleuses of oculomotor nerves
d) Ventral thalamic nucleuses
Pathway continues from thalamus to transverse temporal gyrus as thalamocortical pathway. From here association fibres enters motor areas to provide current spatial information.
Vestibular reflexes
Vestibular apparatus is able to provide reflexes of two types:
a) Postural
b) Statokinetic
Postural reflexes
They are provided during unexpected change of a body position. Its main errand is to prevent a fall. While an individual is standing on a tiltable bed, and an you unexpectedly tilt it, his proximal muscles and deep back muscles will reflectively contract so that certain extremities are extended and the others flexed. Thus the fall is prevented.
Statokinetic reflexes
Are mainly reflective eyes movements, induced by excitation of hair cells in crista ampullaris. Thanks to the existence of the statokinetic reflexes we can stay focus on a certain object while moving a head. This particular reflex is called vestibulo-ocular.
Another important example is a physiological nystagmus. This phenomena has two compounds: fast and slow. Slow compound compensate relative movement of head with regard to an observed object. Fast compound returns eyes to central position. This reflex can be seen while travel by a train and watching trees.
Subchapter Authors: Patrik Maďa and Josef Fontana
![]()