Náplň podkapitoly:
1. Úvod sluchu a vestibulárního systému
2. Sluch
3. Vestibulární systém
_
Úvod sluchu a vestibulárního systému
V této podkapitole si popíšeme dva velmi rozdílné smysly: sluch a vnímání polohy a zrychlení. Tyto smysly jsou často popisovány společně díky úzkým morfologickým vztahům.
_
Sluch
Sluch je smysl, který nám umožňuje vnímat podélné kmitání molekul v okolním prostoru a interpetovat jej jako zvuk. Předpokládá se, že tento smysl původně vznikl k analýze prostředí, především jako nástroj pro registraci blížícího se predátora. Jeho význam se následně rozrůznil mezi jednotlivými taxomy živočichů. Netopýři např. stále spoléhají na sluch jako hlavní analyzátor prostředí a do jisté míry jim nahradili zrak. Jejich sluch je tak citlivý, že odraženou ultrazvukovou vlnou jsou schopni rozlišit drobné nuance v srsti netopýřích samic a samců. U dalších savců pak sluch získal další významnou úlohu. Stal se primárním prostředkem pro registraci zvuků užívaných v komunikaci. Toto je zvlášť zvýrazněno u člověka a kytovců.
Akustika
Akustika je věda zabývající se vznikem a šířením zvukových vln v látkových prostředích. Přítomnost molekul je nutnou podmínkou pro šíření zvuku. Proto uslyšíme přijíždějící lokomotivu, pokud přiložíme ucho ke kolejnici, nebo zvuky šířící se vzduchem. Zvuk se nikdy nemůže šířit ve vakuu. Jak již bylo řečeno, zvuk je podélné kmitání, které se šíří látkovým prostředím. Jelikož se jedná o kmitání podélné, tak prostředí v blízkosti zdroje zvuku se pravidelně zhušťuje a zřeďuje.
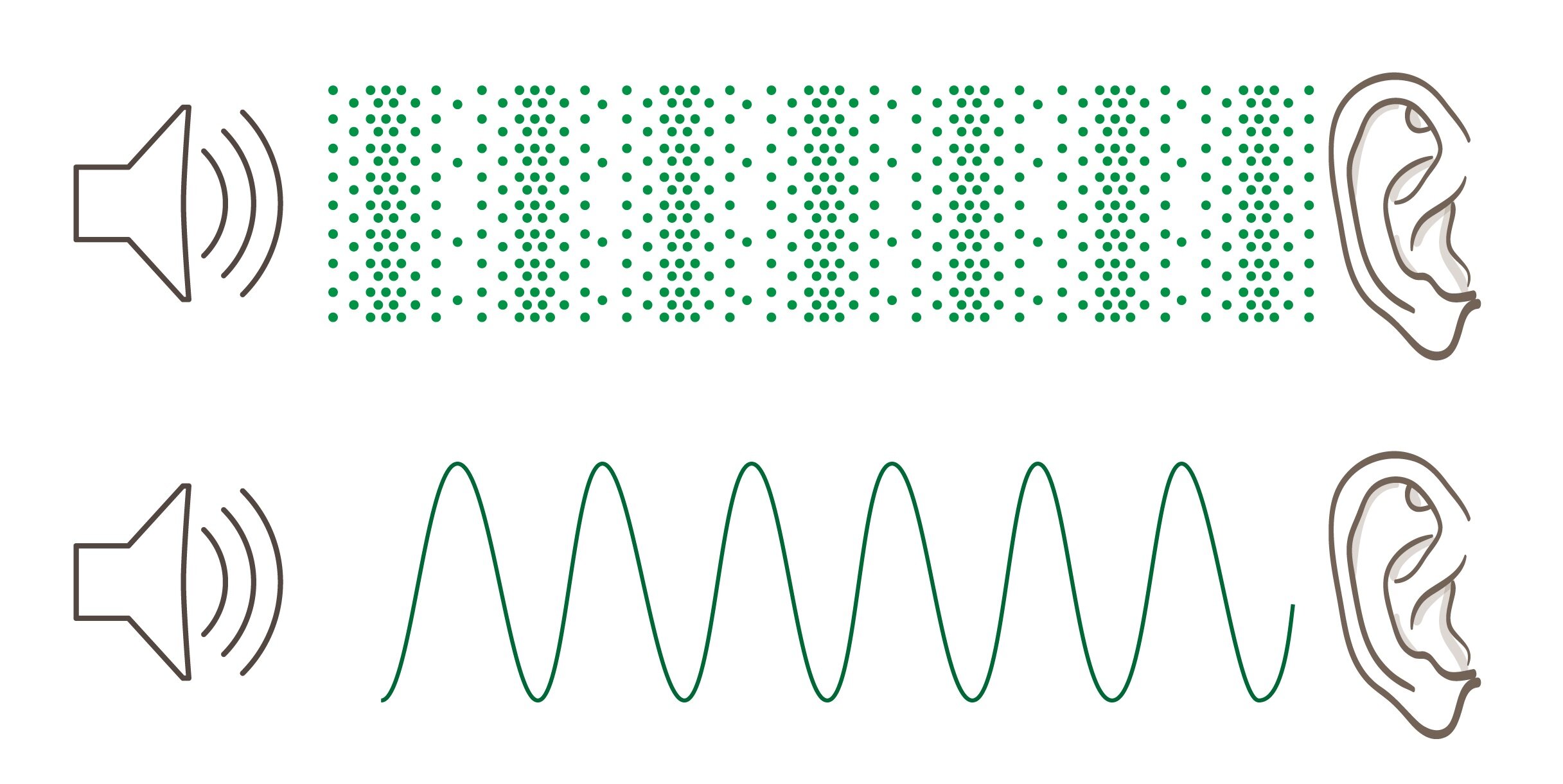
Dvě zhuštění (respektive dvě zředění) prostředí jsou od sebe vzdálená o vlnovou délku. Od zdroje se postupně šíří dále do prostředí rychlostí v, která je při teplotě 0 °C rovna 340 m/s. Lokální změny hustoty vytvářejí kolísavý akustický tlak. Ten je vyšší v místech s vyšší hustotou molekul a naopak klesá ve zředěném látkovém prostředí. Tyto periodické změny v prostředí vytvářejí akustické vlny. Vlastnosti zvukové vlny lze zaznamenat sinusoidou, na níž popisujeme vlnovou délku, amplitudu a frekvenci.
Amplituda, která odpovídá míře zhuštění prostředí, respektive přírůstku akustického tlaku, je subjektivně vnímána jako hlasitost zvuku. Čím vyšší je amplituda daného vlnění, tím je zvuk hlasitější.
Vlnová délka odpovídá výšce tónu. Čím má dané mechanické vlnění vyšší vlnovou délku tím je tón hlubší. Jelikož vlnová délka úzce souvisí s frekvencí (lambda = v/f), tak můžeme říci, že hluboké tóny mají nízké frekvence.
Tón je zvuk, který se skládá jen z vlnění o jediné frekvenci. My se ale v přírodě běžně nesetkáme s jednoduchými tóny, všechny zvuky kolem nás se skládají z více různých podélných vlnění, ať už jde o řeč nebo jen o šumění trávy. V těchto případech mluvíme o harmonických tónech. Harmonický tón se skládá z mnoha vlnění o různé frekvenci. Výšku harmonického tónu určuje vlnění s nejnižší frekvencí, všechna ostatní nabývají tzv. vyšších harmonických frekvencí a určují barvu tónu, díky které například rozeznáme mluvčího. Jako harmonické je označujeme z toho důvodu, že se jedná o celočíselné násobky frekvence základní. Např. frekvence komorního A je 440 Hz. Jeho vyšší harmonické frekvence jsou 880 Hz, 1760 Hz, 3520 Hz, atd. Podle toho, jaké a kolik vyšších harmonických frekvencí je přítomno, rozeznáme např. komorní A zahrané na klavír od toho na housle.
Hluk je směs vlnění o různých frekvencích, které nenabývají vzájemně harmonické frekvence a dokonce se liší svými amplitudami.
To, jaké frekvence vnímáme, je dáno evolučně a velmi se liší mezi jednotlivými druhy. Člověk vnímá zvuky o frekvencích 15 Hz – 16 kHz. Citlivost na vysoké tóny s věkem velmi rychle klesá a u starých lidí je horní hranice jen 5 kHz. Tento jev je velmi snadno pozorovatelný na nástupištích vlakových nádraží. Menší děti si při brždění starých vlakových souprav budou zakrývat uši, jelikož jsou velmi citlivé na vysoké frekvence. Lidé středního věku už většinu vysokých tónů neuslyší a brzdící souprava u nich nevyvolá žádný diskomfort. Lidské ucho je nejcitlivější na tóny o frekvenci v intervalu 2-4 kHz. Tyto frekvence jsou nejvýznamnější pro porozumění mluvené řeči.
Další významným termínem je tzv. sluchový práh. Jedná se o nejmenší akustický tlak, který při dané frekvenci vyvolá sluchový vjem. Je tedy různý pro různé frekvence. Pokud současně s registrovaným tónem znějí i další tóny, zvyšuje se jeho sluchový práh. Tento jev se nazývá maskování.
Hlasitost zvuku je mírou akustického tlaku. Ale jelikož akustické tlaky jsou velmi malé a špatně se s nimi pracuje, byla arbitrárně stanovena veličina hladina intenzity zvuku Lp (někdy též hladina akustického tlaku). Její jednotkou je decibel – dB. Pro její vyjádření platí následující vztah:
Lp = 20 log(Px/P0)
Px – aktuální akustický tlak
P0 – sluchový práh pro danou frekvenci
Ze vztahu vyplývá, že pokud se aktuální akustický tlak zvýší tisíckrát, tak hladina intenzity zvuku stoupne o 60 dB. Jelikož se argument logaritmu zvýšil tisíckrát, stačí nám znát, že dekadický logaritmus 1000 je 3. Hladina intenzity zvuku pak narostla o Lp = 20 x 3. Zvýšila se tedy o 60 dB.
Pro stanovení vlastní hlasitosti zvuku, pak potřebujeme naměřit hladinu intenzity pro danou frekvenci a zanést ji do grafu. To musíme provést pro různé frekvence, které daný jedinec slyší subjektivně stejně hlasitě. Vznikne nám křivka, kterou označujeme jako izofona. Jinak řečeno, izofona stanoví pro každý tón o určité frekvenci takovou hladinu intenzity zvuku, při které jsou tyto tóny stejně hlasité. Izofona, jež spojuje nejmenší hladiny intenzity zvuku, při kterých vlnění o dané frekvenci vyvolá sluchový vjem, se nazývá práh slyšitelnosti. Práh slyšitelnosti je tedy křivka, která spojuje sluchové prahy pro jednotlivé tóny. Pokud budeme u jednotlivých tónů dále navyšovat hladinu intenzity, dostaneme se časem do hodnot, které nevnímáme jako zvuk, ale jako bolest. Propojením těchto hodnot izofonou získáme práh bolesti.
Při frekvencích kolem 2000 Hz odpovídají hladiny intenzity následujícím jevům:
4 dB – Práh slyšitelnosti
20 dB – Hluboké ticho (např. v akustickém studiu)
30 dB – Šepot
60 dB – Běžný hovor
100 dB – Koncert
130 dB – Práh bolesti
140 dB – Akustické trauma (v blízkosti startujícího proudového letadla)
Hlasitost zvuku se měří v klinické praxi při tzv. subjektivní audiometrii. Zde se udává ve fonech. Při běžném frekvenčním rozsahu (2-4 kHz) odpovídá jeden fon přibližně jednomu decibelu. Běžný hovor má hlasitost 60 fonů. Další možností vyjádření hlasitosti jsou sony. V sonech udáváme kolikrát slyšíme daný tón hlasitěji oproti stanovenému základu. Definice říká, že 1 son je hlasitost tónu o hladině intenzity 40 dB při frekvenci 1000 Hz. 2 sony má takový tón, který subjektivně vnímáme dvakrát hlasitěji.
Percepce zvuku
Percepcí zvuku se rozumí přenos mechanického podélného vlnění přes sluchový aparát, jeho kódování ve vnitřním uchu na Cortiho orgánu a konečně vlastní interpretace ve vyšších etážích CNS.
Zevní ucho
Rolí zevního ucha je zachytit mechanické vlnění okolního vzduchu a vnějším zvukovodem jej přivést až na bubínek (membrana tympani). Všimněte si, že jeho úloha spočívá v zachycení zvuku okolí, zachycuje tzv. vzdušné vedení zvuku. Zevní ucho se neuplatňuje při poslechu vlastního hlasu, který se šíří v kostech lebky po rozvibrování dutin a přímo se přenáší na perilymfu vnitřního ucha. Tento jev nazýváme kostní vedení.
Střední ucho
Střední ucho je laterálně ohraničeno bubínkem, tedy pružnou membránou, která se vlivem akustického tlaku rozkmitá s frekvencí a amplitudou odpovídající danému zvuku. Kmitání bubínku se přenáší na kladívko, které spojeno s kovadlinkou a ta konečně s třmínkem. Baze třmínku je zavzata do foramen ovale a jeho pohyb rozkmitává perilymfu. Souhrnně se jedná o důmyslný systém pák, které zabraňují ztrátám akustické energie. Respektive mění vlastnosti rozhraní vzduch a perilymfa.
Pokud by mezi vzduch a perilymfu nebylo vřazeno střední ucho, došlo by na tomto rozhraní k částečnému odrazu zvuku, který by velmi komplikoval jeho percepci. Střední ucho tak tvoří převodní aparát, který významně redukuje ztráty, jelikož plocha bubínku je násobně větší než baze třmínku. Uvažujme, že součin tlaku a plochy je konstantní, pak relativně malý akustický tlak na bubínek je pomocí kůstek středního ucha převeden do perilymfy jako násobek akustického tlaku působícího na membrana tympani a poměru ploch bubínku a foramen ovale. Jednoduše řečeno soustava kůstek funguje jako zesilovač akustického tlaku.
Na středoušní kůstky se dále upínají dva svaly: m. stapedius a m. tensor tympani. Oba se reflexně stahují při nadměrném zvukovém stimulu. V kontrahovaném stavu fungují jako tlumiče vibrací a snižují tím zesilovací efekt soustavy středoušních kůstek. Tak ochraňují citlivý aparát ve vnitřním uchu před excesivním zvukovým stimulem.
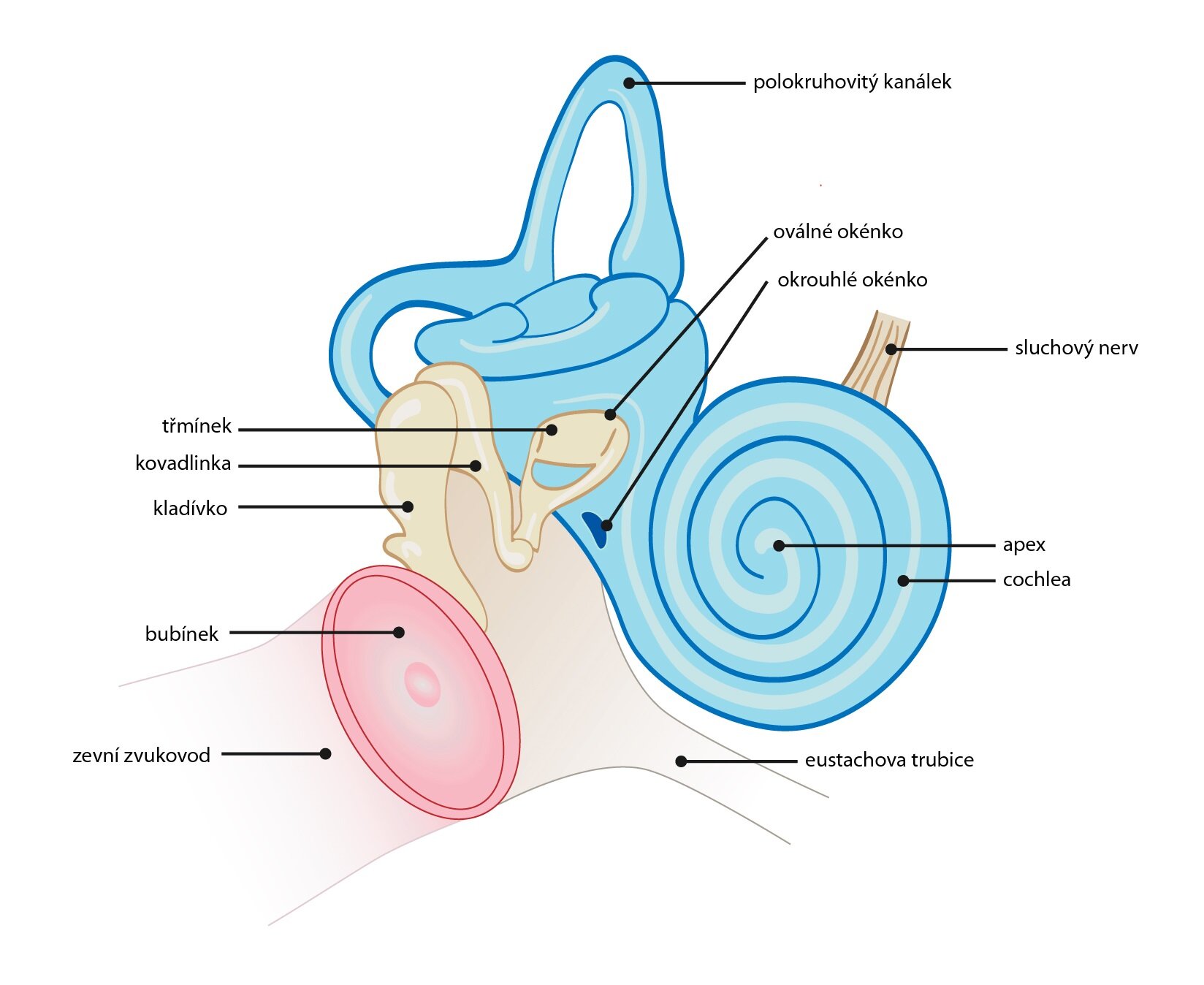
Vnitřní ucho
Vnitřní ucho se nachází v dutině temporální kosti zvané kostěný labyrint. V něm nacházíme jak vestibulární systém, tak kochleu, která obsahuje vlastní receptory sluchu. Kochlea je hlemýžďovitě stočený útvar se třemi dutinami, které se táhnou celou délkou hlemýždě: scala vestibuli, scala media a scala tympani.
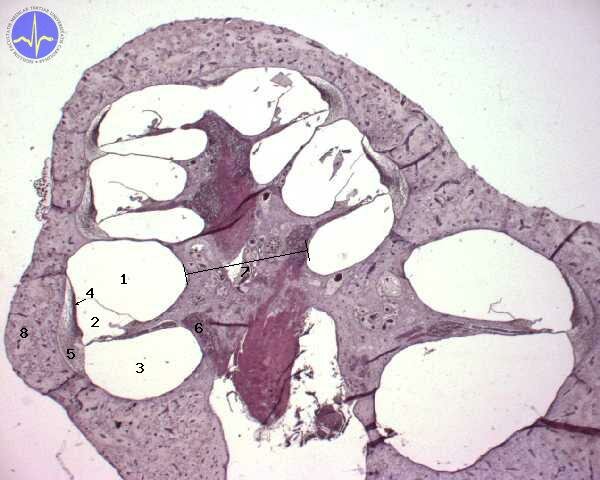
1 – Scala vestibuli, 2 – Scala media, 3 – Scala tympani, 4 – Stria vascularis, 5 – Ligamentum spirale, 6 – Ganglion spirale, 7 – Modiolus, 8 – Kostěný hlemýžď
1) Scala vestibuli naléhá na foramen ovale. Obsahuje perilymfu a táhne se na vnitřním povrchu závitů.
2) Scala media obsahuje vlastní receptory sluchu. Jedná se o vláskové buňky, které jsou usazeny na bazilární membráně Cortiho orgánu. Omývá je endolymfa – výplň scala media.
3) Scala tympani je vyplněna perilymfou. V délce hlemýždě se táhne po vnější straně závitů, naléhá na foramen rotundum.
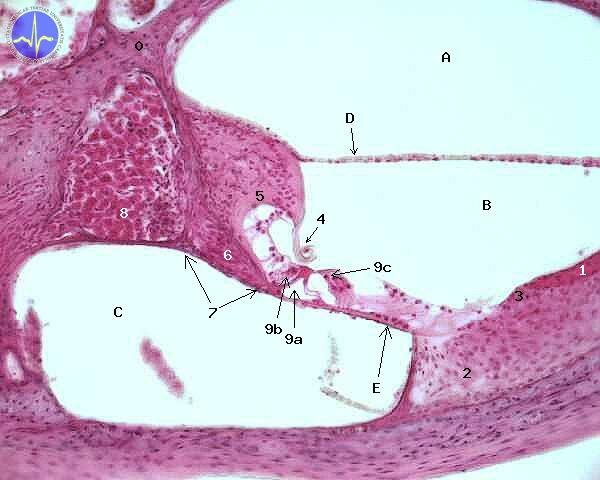
0 – Modiolus, 1 – Stria vascularis, 2 – Ligamentum spirale, 3 – Prominentia spiralis, 4 – Membrana tectoria, 5 – Limbus spiralis, 6 – Vlákna nervus cochlearis, 7 – Kostěná lamina spiralis, 8 – Ganglion spirale, 9a – Vnitřní Cortiho tunel, 9b – Vnitřní vlásková buňka, 9c – Vnější vláskové buňky, A – Scala vestibuli, B – Scala media, C – Scala tympani, D – Membrana vestibularis, E – Membrana basilaris
Jednotlivé dutiny jsou vzájemně odděleny membránami. Mezi scala media a scala tympani se jedná o již zmíněnou bazilární membránu Cortiho orgánu. Scala vestibuli je pak od scala media oddělena Reissnerovou membránou, ze které se odděluje tektoriální membrána. Ta je v přímém kontaktu s apikálním povrchem vláskových buněk.
Vznik akčního potenciálu vláskové buňky
Pokud se rozechvěje baze třmínku, dojde k přenosu vibrací do perilymfy scala vestibuli. Tím se začne pohybovat i Reissnerova membrána a s ní související membrána tektoriální. Vláskové buňky mají na svém apikálním povrchu stereocílie. Tření stereocílií o tektoriální membránu mění potenciál membrány vláskových buněk. To je vnímáno nervovými zakončeními prvního neuronu sluchové dráhy. Vibrace tektoriální a Reissnerovy membrány se samozřejmě rozšíří i na endolymfu a bazilární membránu Cortiho orgánu. K registraci zvuků tak ve skutečnosti přispívá chvění celé soustavy.
Stereocílie jsou v přímém kontaktu s endolymfou, jejíž složení se podobá intracelulární tekutině. Endolymfa obsahuje vysokou koncentraci draslíkových iontů (okolo 150 mmol/l), které jsou do ní secernovány draslíkovými kanály z buněk stria vascularis. Do buněk stria vascularis se ionty dostávají pomocí stejného přenašeče, jaký je i v ascendentním raménku Henleovy kličky tj. Na+/K+/2Cl– symportem. Při správném vychýlení stereocílie se v její membráně otevírá kanál, který propouští K+ dovnitř vláskové buňky. Jedná se tedy o iontový kanál, který je otevírán mechanicky. K+ se pohybuje po směru elektrického gradientu a jeho influx vede k depolarizaci membrány. Z vláskové buňky je K+ následně vypuštěn do perilymfy, kde je fyziologicky nízká koncentrace K+ (jako v extracelulární tekutině). K+ se tedy pohybuje po svém koncentračním gradientu a jeho exfulx vede k repolarizaci. Aby se celý proces uzavřel, jsou K+ ionty aktivně přečerpávány z perilymfy zpět do endolymfy, odkud mohou opět vstoupit do stereocílie.
Klinické korelace:
Zvýšení permeability mezi prostory endolymfy a perilymfy je jedním z mechanismů zodpovědných za vznik Meniérovy choroby, pro níž jsou typické záchvaty zhoršeného sluchu a závratí.
Genetický defekt K+ kanálu v membráně buněk stria vaskularis vede k hluchotě a dále k syndromu dlouhého QT intervalu. Kanál je totiž také exprimován v srdci, kde se účastní repolarizace. Jeho defekt tedy vede k prodloužení repolarizace, což se na EKG projevuje prodloužením QT intevalu.
Kódování frekvence a intenzity zvuku
Vibrace se v perilymfě šíří od formen ovale ke středu hlemýždě (helikotromě). Aby mohly být rozlišeny jednotlivé tóny, je bazilární membrána modifikovaná tak, že se směrem k helikotremě postupně zmenšuje její tuhost. To má dva fyzikální důsledky: pro danou frekvenci se zvyšuje amplituda, se kterou se bazilární membrána chvěje a zároveň se snižuje rychlost šíření vibrací. Díky změnám tuhosti – respektive akustických vlastností – v průběhu bazilární membrány existuje pro každou frekvenci úsek stěny, kde dochází k rezonanci vibrací. V tomto úseku bazilární membrány jsou amplitudy obzvláště vysoké a jsou zde registrovány vláskovými buňkami, jako tzv. kmitající maximum. Tato teorie se nazývá fonotopie.
V závislosti na úseku bazilární membrány (jednotlivé úseky se liší kmitajícím maximem), na kterém se vláskové buňky nacházejí, je lze rozdělit do skupin. Každá skupina je citlivá na jiné frekvence vibrací. Vysoké frekvence mají svá kmitající maxima při foramen ovale. Nízké frekvence jsou naopak registrovány u helikotremy. Nejvíce skupin vláskových buněk se nachází v intervalu 2-4 kHz. Zde dokáže lidské ucho rozlišit tón lišící se o pouhé 3 Hz.
Sluchová dráha
Sluchová dráha je obvykle popisována jako čtyřneuronová. V nedávné době však byly zjištěny v průběhu dráhy další synapse a její popis se tedy zkomplikoval.
Vláskové buňky nemají vlastní axony, proto prvním neuron je až bipolární buňka, jejíž perikaryon se nachází v ggl. spirale cochleae. Jeho dendrit tvoří synapsi s vláskovou buňkou a tvoří kochleární část n. vestibulocochlearis. Spolu s ním tak vstupuje do ncc. cochleares v kaudální části pons Varoli. V kochleárních jádrech se přepojuje na druhý neuron. Z ncc. cochleares vystupují tři svazky:
1) Z nc. cochlearis ventralis vychází corpus trapezoideum
2) Z nc. cochlearis ventralis vychází stria acustica intermedia
3) Z nc. cochlearis dorsalis vychází stria acustica dorsalis
Většina neuronů všech tří svazků se kříží a na kontralaterální straně společně formují lemniscus lateralis. Směřují rostrálně a přepojují se na třetí neuron v colliculi inferiores mesencephala. Již zde dochází k rozporu se čtyřneuronovou teorií, neboť některá vlákna se přepojí již v nc. olivaris superior, jiná v nc. corporis trapezoidei a jiná konečně v nc. lemnisci lateralis.
Z colliculi inferiores se axony třetího neuronu táhnou až do thalamu, respektive jeho corpus geniculatum mediale na ipsilaterální straně, a zde se přepojují na čtvrtý neuron. Čtvrté thalamické neurony vytvářejí svými axony svazek zvaný radiatio acustica (popisně tr. geniculocorticalis). Ten vstupuje do primární sluchové kůry, kterou nacházíme na g. temporalis superior na tzv. Heschlových závitech.
Primární sluchová kůra je stejně jako celý průběh sluchové dráhy uspořádána tonotopicky. To znamená, že určité frekvence aktivují pouze určitá vlákna a následně určitá místa v kůře. Probíhá zde identifikace jednotlivých tónu, analýza fonémů a konečně také přibližné určení místa zdroje zvuku.
Z primární sluchové oblasti pak vedou asociační vlákna do sekundární sluchové oblasti. Zde se vytvoří vnitřní obraz registrovaného zvuku, který pak může být převeden do asociačních oblastí, kde je mu přiřazen přesný význam. Má se za to, že v sekundární sluchové kůře sídlí i zvuková paměť, díky které rozeznáváme známé zvuky.
Je dobré zdůraznit, že jednotlivé etáže sluchové dráhy často odevzdávají část vláken do kontralaterální dráhy. Proto má každé ucho bilaterální projekci. Jednostranná léze tedy nezpůsobí vážnou poruchu sluchu, pouze ztěžuje diskriminaci místa zdroje zvuku.
_
Vestibulární systém
Vestibulární aparát je soustavou dvou čidel: statického a kinetického. Proto je také někdy nazýván statokinetickým aparátem. Stejně jako kochlea je zavzat do labyrintu v kosti skalní. Statické čidlo tvoří utrikulus a sakulus. Kinetické čidlo je tvořeno třemi na sebe vzájemně kolmými polokruhovitými kanálky.
Všechny součásti vestibulárního systému jsou omývány perilymfou a jsou vyplněny endolymfou. Obě tekutiny mají složení, které bylo popsáno výše u vnitřního ucha.
Statické čidlo
V utrikulu a sakulu se nacházejí vyvýšeniny epitelu zvané makula. Na jejich povrchu se nacházejí vláskovité buňky se stereocíliemi. Makula v utrikulu se nachází kolmo na osu těla. Makula v sakulu leží v ose těla. Jejich povrch je pokryt obalem z mukopolysacharidů (kupola), který obsahuje otolity. Jedná se o útvary z krystalů kalcitu (CaCO3). Pokud je hlava ve vzpřímené poloze, ohýbají otolity stereocílie vláskových buněk pouze makulu v sakulu. Při úklonu hlavy na stranu se vlivem gravitace přitisknou otolity utrikulu k jeho makule a podráždí stereocílie. Jejich podráždění probíhá stejným mechanismem jako těch v Cortiho orgánu. Statické čidlo tak reaguje na gravitační zrychlení a v každém okamžiku informuje CNS o poloze hlavy v prostoru oproti gravitačnímu poli. Statické čidlo dále reaguje na lineární zrychlení. Pokud je vektor zrychlení vertikální, aktivují se makula v sakulu (např. jízda výtahem). Pokud se naopak vektor zrychlení nachází v horizontální rovině aktivují se macula utriculi (např. zrychlování při jízdě v autě).
Je nutné si uvědomit, že akční potenciály jsou vysílány ve velkých frekvencích zejména při přesunu otolitů – tedy při změnách polohy hlavy. Pokud je poloha hlavy konstantní, tak se otolity nepohybují a nedojde k podráždění stereocílií a vzniku akčního potenciálu. CNS pak pracuje s poslední známou informací o poloze v prostoru. Ta je při sebemenší změně upravena na novou polohu.
Kinetické čidlo
Kinetické čidlo se skládá ze tří polokruhovitých kanálků: canalis semicircularis anterior, posteriol et lateralis. V každém z kanálků se nachází crista ampularis, která má na svém povrchu vláskové buňky se stereocíliemi. Povrch crista ampularis je – stejně jako makula v sakulu a utrikulu – pokryt rosolovitým mukopolysacharidovým obalem s otolity. V případě změny polohy hlavy se otolity přesunou a podráždí stereocílie.
Kinetické čidlo reaguje na úhlové zrychlení. V souladu s prvním Newtonovým zákonem (zákon setrvačnosti) si endolymfa snaží uchovat svou polohu i při rotačních pohybech hlavou. Vzhledem k polokruhovitým kanálkům se tak pohybuje proti směru otáčení a mění polohu rosolovité kupoly. Tím aktivuje stereocílie.
Vestibulární dráha
Vestibulární dráha je považována za tříneuronovou dráhu. Stejně jako vláskovité buňky Cortiho orgánu nemají senzorické buňky makul a crista ampularis vlastní axony. Jsou inervovány neurony ggl. vestibulare, které tvoří vestibulární část n. vestibulocochlearis (vzniká ještě ve vnitřním uchu). Některé axony prvních neuronů nepokračují dále ve vestibulární dráze, ale tvoří tr. vestibulocerebelaris directus a vstupují přímo do mozečku, který tak má aktuální informace o poloze a zrychlení dříve než kůra. Zbylé axony prvního neuronu vstupují do pons Varoli, kde se nacházejí ncc. venstibulares pontis. Jsou celkem čtyři: nc. lateralis, nc. superior, nc. medialis a nc. inferior. Zde dojde k přepojení na druhý neuron. Tato jádra přijímají i informace z hlubokých mechanoceptorů. Odtud dojde k přepojení do různých struktur:
a) Do mozečku, jako tr. vestibulocerebelaris indirectus – tato dráha poskytuje mozečku informaci o poloze zpřesněnou o hluboké čití
b) Do míchy, jako tr. vestibulospinalis – tak mohou vyvolávat posturální reflexy
c) Do jader okohybných nervů – zde zprostředkují statokinetické reflexy
d) Do thalamu na další zpracování, zejména do ncc. ventrales thalami
Některá vlákna postupují i přímo do kůry nebo do retikulární formace.
K přepojení na třetí neuron dochází v thalamu v jeho ncc. ventrales thalami. Odtud pokračuje thalamokortikální dráhou do g. temporalis transversus, kde se nachází projekce vestibulárního aparátu. Ta vysílá asociační vlákna jednak do asociačních oblastí pro vědomé zpracování polohy, ale také do motorických oblastí (zejména sekundární), kde se podílí na plánování pohybů.
Vestibulární reflexy
Vestibulární aparát je schopen při adekvátním podnětu vyvolávat reflexy, které lze rozdělit do dvou typů:
a) Posturální
b) Statokinetické
Posturální reflexy
Zvané také vzpřimovací nebo postojové. Jsou vyvolány při náhlé změně polohy vyvolané vnějšími vlivy a jejich úkolem je předejít pádu jedince. Uveďme si experimentální příklad. Představte si, že požádáme jedince, aby zaujal polohu na všech čtyřech na pohyblivé podložce a tu prudce na jednu stranu nakloníme. Reflexně dojde k aktivaci proximálních svalů končetin a hlubokých svalů zad tak, aby se flektovaly končetiny na jedné straně a naopak extendovaly končetiny v místě poklesu podložky. Tím dojde k částečnému (nebo úplnému) vyrušení změny polohy vyvolané vnějším vlivem.
Statokinetické reflexy
Statokinetické reflexy se objevují při pohybu otolitů v polokruhovitých kanálcích. Zvláštní význam mají zejména reflexní pohyby očí, které nám umožňují mít zaostřeno na daný objekt i přes pohyby hlavy. Když hlava rotuje doleva, oči se reflexně a úměrně přesunou doprava, tak nedojde k rozostření obrazu. Tento reflex se nazývá vestibulo-okulární.
Dalším příkladem statokinetického reflexu je fyziologický nystagmus. Ten má dvě složky – pomalou a rychlou. Pomalá složka je pohyb očí, který kompenzuje relativní pohyb hlavy oproti sledovanému objektu. Rychlá složka je pohyb očí, který je vybaven vestibulárními centry v mozkovém kmeni a staví oči zpět do středního postavení. Tento reflex je dobře pozorovatelný při jízdě vlakem, když sledujete podél kolejí se míjející stromy. Jak se vlak pohybuje, oči stále následují sledovaný strom (pomalá složka). Jakmile zmizí ze zorného pole, oči se v rámci milisekund vrátí do výchozího postavení (rychlá složka) a vyhledají další strom. Celý cyklus se pak opakuje.
Autoři podkapitoly: Patrik Maďa a Josef Fontana
![]()