Content:
1. Structure of the cell
_
Structure of the cell
The cell is basic structural unit of eukaryotes. Its surface is covered by plasma membrane separating the cellular content from outer environment. This membrane creates barrier which controls substances that enter or leave the cell. It is composed of phospholipid bilayer with embedded cholesterol molecules, proteins and oligosaccharides. The hydrophobic fatty-acid chains face each other and their hydrophilic polar heads are oriented outside and form both surfaces of the membrane. Cell can be divided into two major compartments – the nucleus and the cytoplasm containing organelles.
Nucleus
The nucleus imitates the shape of cell but most often is rounded and its size varies in the range 5 to 10 µm diameter. It consists of nuclear envelope, chromatin, nucleolus and nucleoplasm.
Nuclear envelope
Nuclear envelope is formed by two membranes surrounding nucleus. The inner membrane is proper nuclear membrane and the outer one is continuous with endoplasmic reticulum.
Chromatin
Chromatin is the second component of the nucleus. Two types of chromatin are distinguished: heterochromatin – dark staining in light microscope, and euchromatin – lightly staining form. Heterochromatin represents highly dense (spiralized) genetic material and euchromatin appears more dispersed. Chromatin is formed by coiled DNA molecules bound to histones (basic proteins). DNA molecules contain most of genetic informations.
Nucleolus
Nucleolus is round-shaped structure about 1 μm in diameter localized in protein-synthesizing cells.
Nucleoplasm
Nucleoplasm fills the space between chromatin. It consists of proteins, various metabolites and ions.
Cytoplasm
Cytoplasm contains many important specialized organelles necessary for viability of cell. Membranous organelles include: mitochondria, endoplasmic reticulum, Golgi apparatus, lysosomes, peroxisomes and secretory granules.
Mitochondria
Mitochondria are semiautonomous organelles which do not need cellular nucleus for their division. Mitochondria possess outer and inner mitochondrial membranes differing in many characteristics.
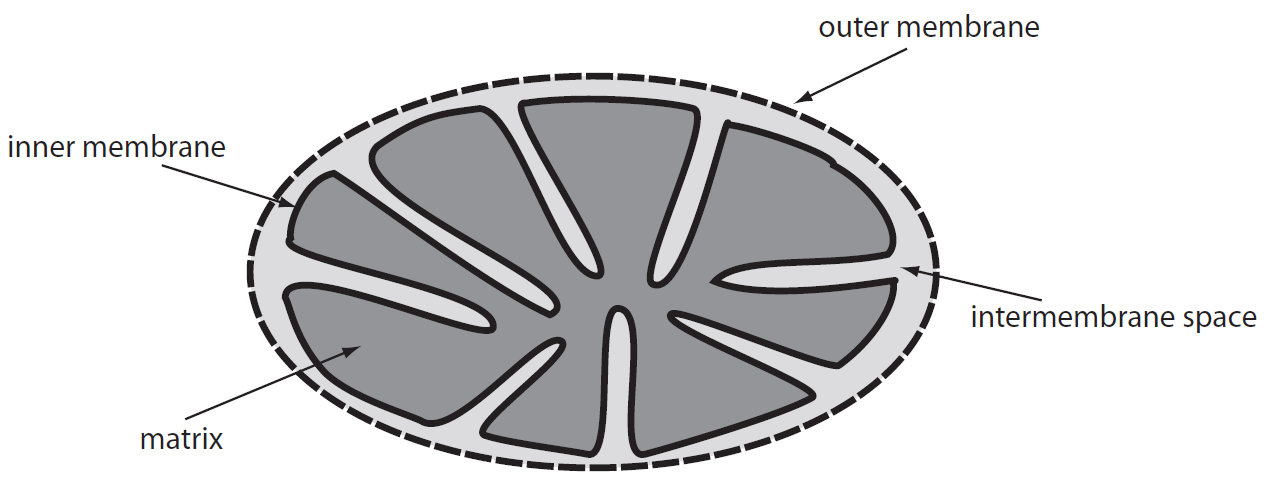
The outer membrane is smooth and permeable for many molecules as large as 10 kDa. It also contains cholesterol. The inner membrane is permeable only for small molecules in range 100-150 Da. It contains smaller amount of cholesterol and it is arranged into numerous mitochondrial cristae which increase the inner membrane surface. Space between these two membranes is connected to space between mitochondrial cristae. The inner mitochondrial membrane surrounds the space called mitochondrial matrix. Some mitochondria form tubular cristae – these mitochondria are present in cells involved in steroid metabolism (adrenal cortex, genital glands). The inner membrane contains proteins performing metabolic reactions (e.g. cytochromes, enzymes of the Krebs cycle). One of the main functions of mitochondria is ATP production (macroergic compound). ATP generated in the inner membrane is transported into the cytoplasm where is used as energy for metabolic processes (membranous transport of substances, mechanical muscle contraction, synthesis reaction).
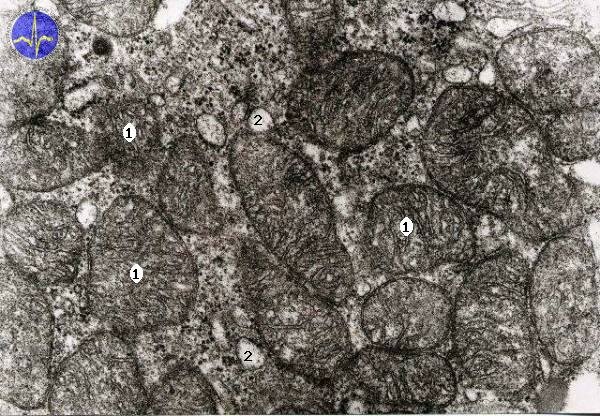
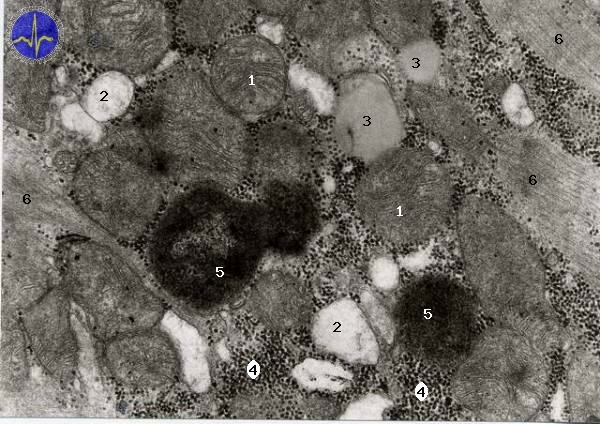
Mitochondria with cristae
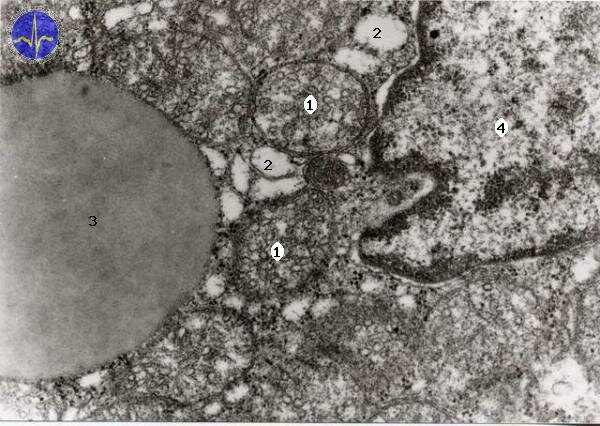
1 – Tubular mitochondria, 2 – Smooth endoplasmatic reticulum, 3 – Lipid droplet, 4 – Nucleus
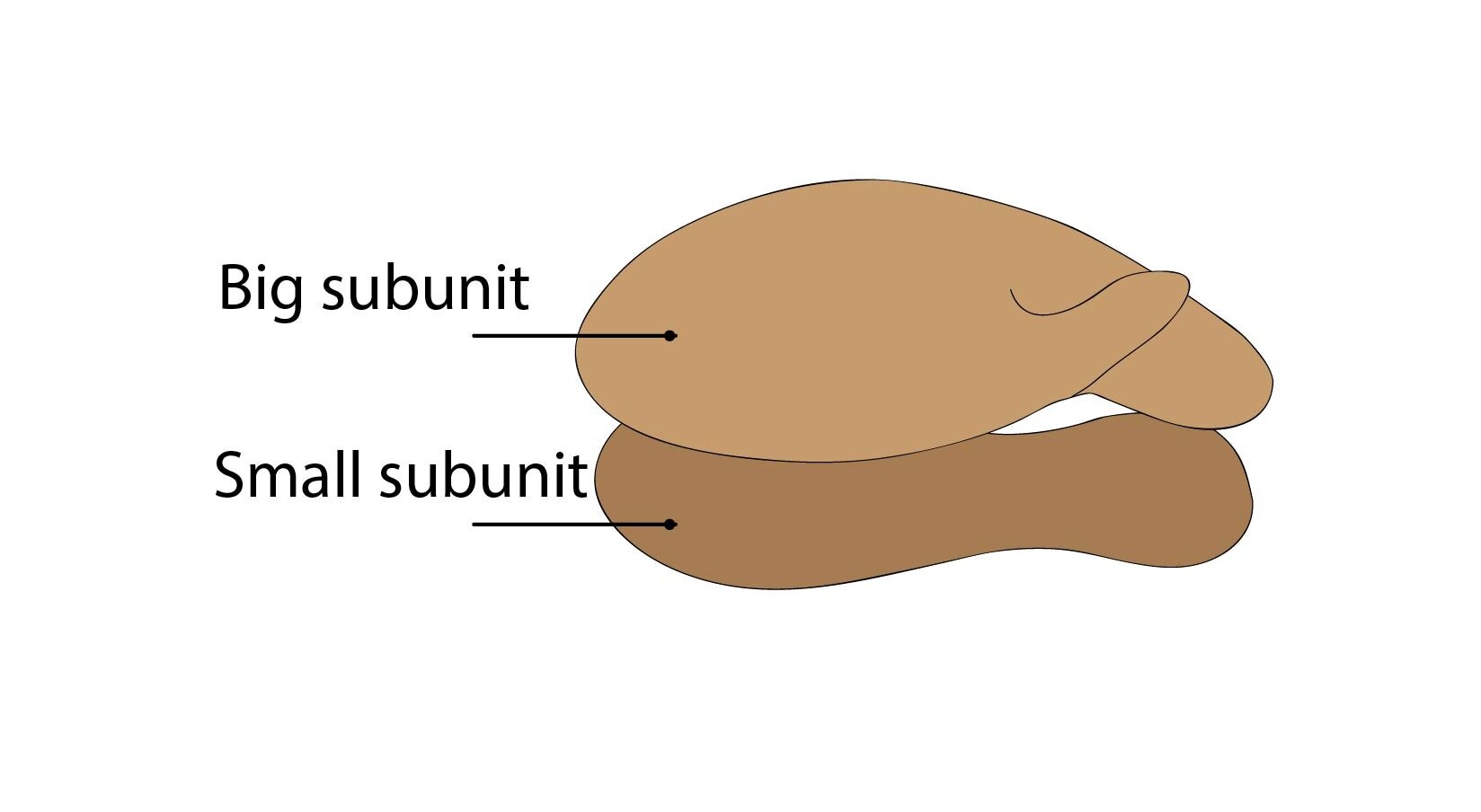
Ribosomes
Ribosomes are granular organelles ensuring proteosynthesis. They are composed of small and large subunit dispersed in cytoplasm and after catching by mRNA (messenger RNA), these subunits merge together to form ribosome. After translation are these subunits divided again. Ribosomes can be found free in cytoplasm or attached to ribosomal membrane (forming rough endoplasmic reticulum).
Endoplasmic reticulum (ER)
Endoplasmic reticulum is a group of flattened elongated sacs surrounded by one-layer membrane performing synthesis of necessary molecules. Because of connection between sacs is this organelle called reticulum. ER is present in cell in two different forms: smooth endoplasmic reticulum and rough endoplasmic reticulum. The rough ER contains free ribosomes and is responsible for proteosynthesis. The smooth endoplasmic reticulum ensures steroid and lipid synthesis. The special type of smooth endoplasmic reticulum is sarcoplasmic reticulum contained in smooth muscle cells involved in regulation of Ca2+ concentration.
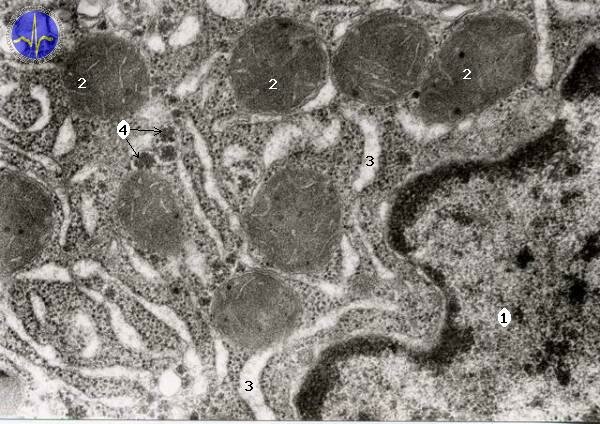
1 – Nucleus, 2 – Mitochondria, 3 – Endoplasmatic reticulum, 4 – Polyribosomes
Golgi apparatus (GA)
Golgi apparatus is a complex of cisternae and sacs dispersed in cytoplasm. It creates functional border between cytoplasm and endoplasmic reticulum. Ensures modification of proteins produced in ER – glycosylation, phosphorylation etc. It also participates in creation of secretory granules in cytoplasm – these granules are concentrated products of endoplasmic reticulum.
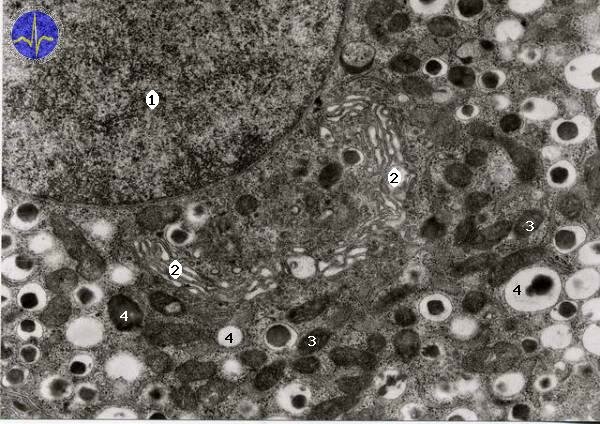
1 – Nucleus, 2 – Golgi apparatus, 3 – Mitochondria, 4 – Transporting vesicles
Secretory granules
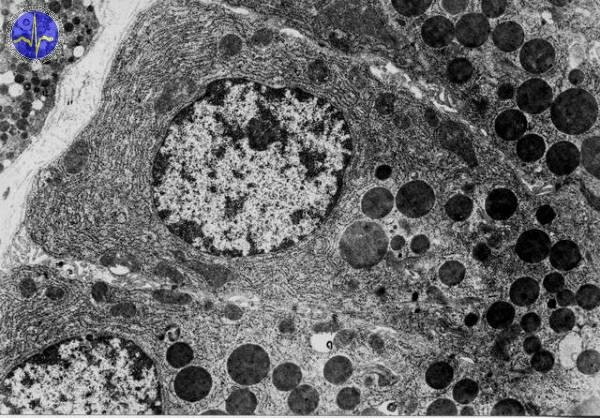
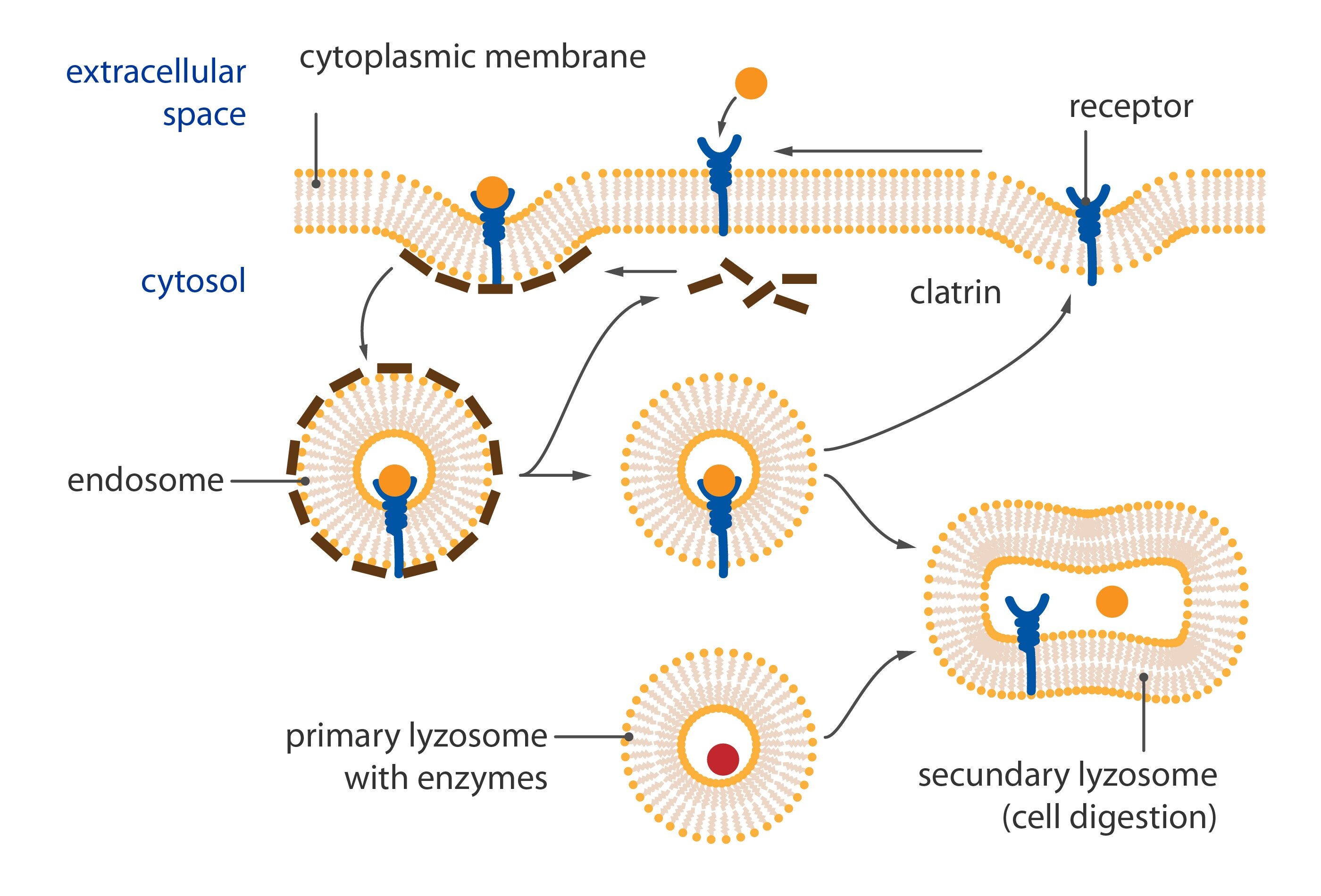
Lysosomes
Lysosomes are hollow rounded organelles containing enzymes e.g. alkaline phosphatase, proteases, lipases etc. We can distinguish two different types of lysosomes: primary lysosomes containing enzymes produced from ER and then modified in Golgi apparatus which finally become the primary lysosomes. The secondary lysosomes are created by merging of primary lysosome and phagosome. Secondary lysosomes are larger than primary and can be distinguished into autolysosomes and heterolysosomes. Autolysosomes are involved in degradation of cellular particles and heterolysosomes (heterophagosomes) eliminate phagocytized material. Tercial lysosomes contain residual bodies (e.g. lipofuscin granules typically yellowish in light microscope).
1 – Mitochondria, 2 – ER, 3 – Lipid droplets, 4 – Glykogen inclusions, 5 – Lipofuscin inclusions, 6 – Myofibrils
Peroxisomes
Peroxisomes are small organelles enclosed by membrane. They are produced by endoplasmic reticulum and are responsible for protection of cell against oxygen free radicals and production of bile acids. They are also involved in lipid metabolism.
Cytoskeleton
The cellular cytoskeleton includes: microfilaments (actin filaments), intermediate filaments and microtubules. These protein structures are responsible for mechanical support of cell and they participate in cell division or transport of various substances.
Microfilaments (actin filaments)
Microfilaments are fibrillar structures about 7 nm in diameter composed of actin molecules. In most of the cases, these fibers form a meshwork just beneath the plasma membrane, but other structural organization can be also present. Microfilaments participate in endocytosis, exocytosis, contraction of microvilli, cellular movements and flow of cytoplasmic components. They also ensure the contractility of whole body – muscle cells.
Intermediate filaments
Intermediate filaments are about 10 to 12 nm thick and they represent the second cytoskeleton type. They can be distinguished using immunohistochemistry because they exhibit different proteins. This group includes cytokeratins contained in epithelial cells, desmin contained in smooth muscle cells, vimentin with mesenchymal origin found in embryonal cells, neurofilaments contained in most of neurons, glial fibrillary acidic protein in astrocytes and lamins A, B, C.
Microtubules
Microtubules measure about 25 nm in diameter and are formed by alpha- and beta-tubulin molecules. Tubulin creates protofilaments arranged in the wall of microtubule in number of 13. Microtubules ensure cellular shape, the movement of chromosomes along the spindle apparatus, or vesicular movement (transport of substances) in cell. Function of microtubules is inhibited by colchicine and vinblastine – they are toxic and stop the cell division.
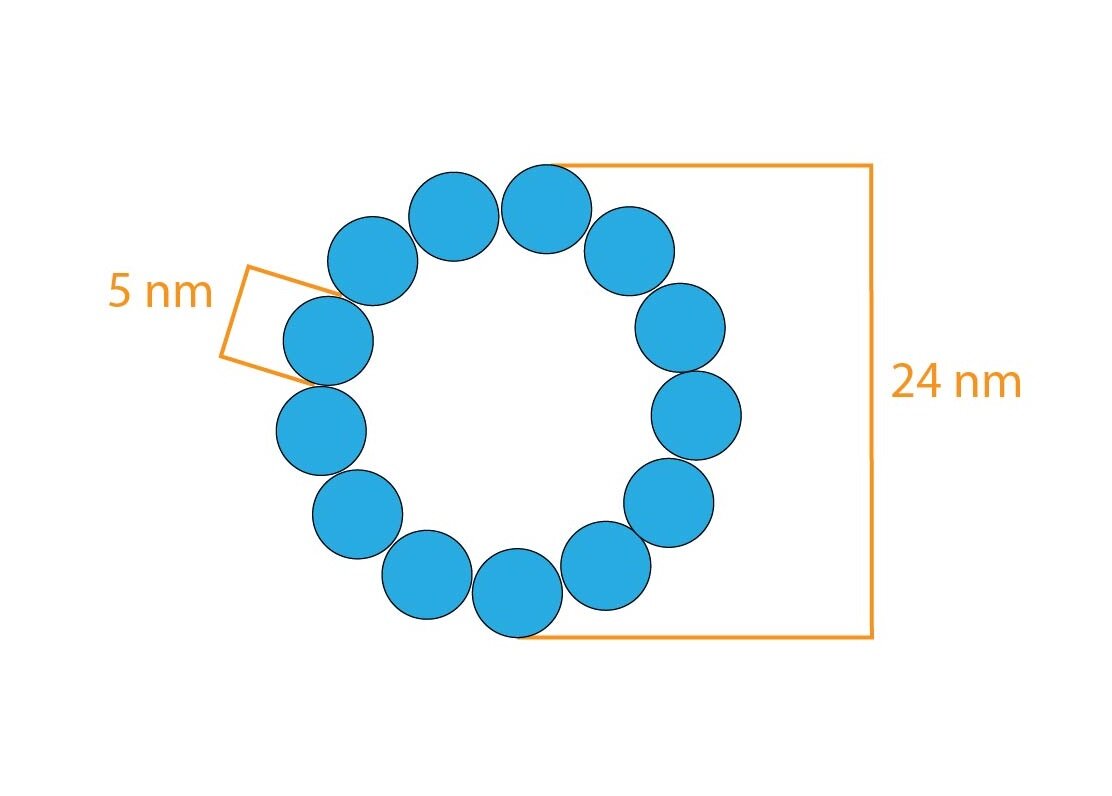
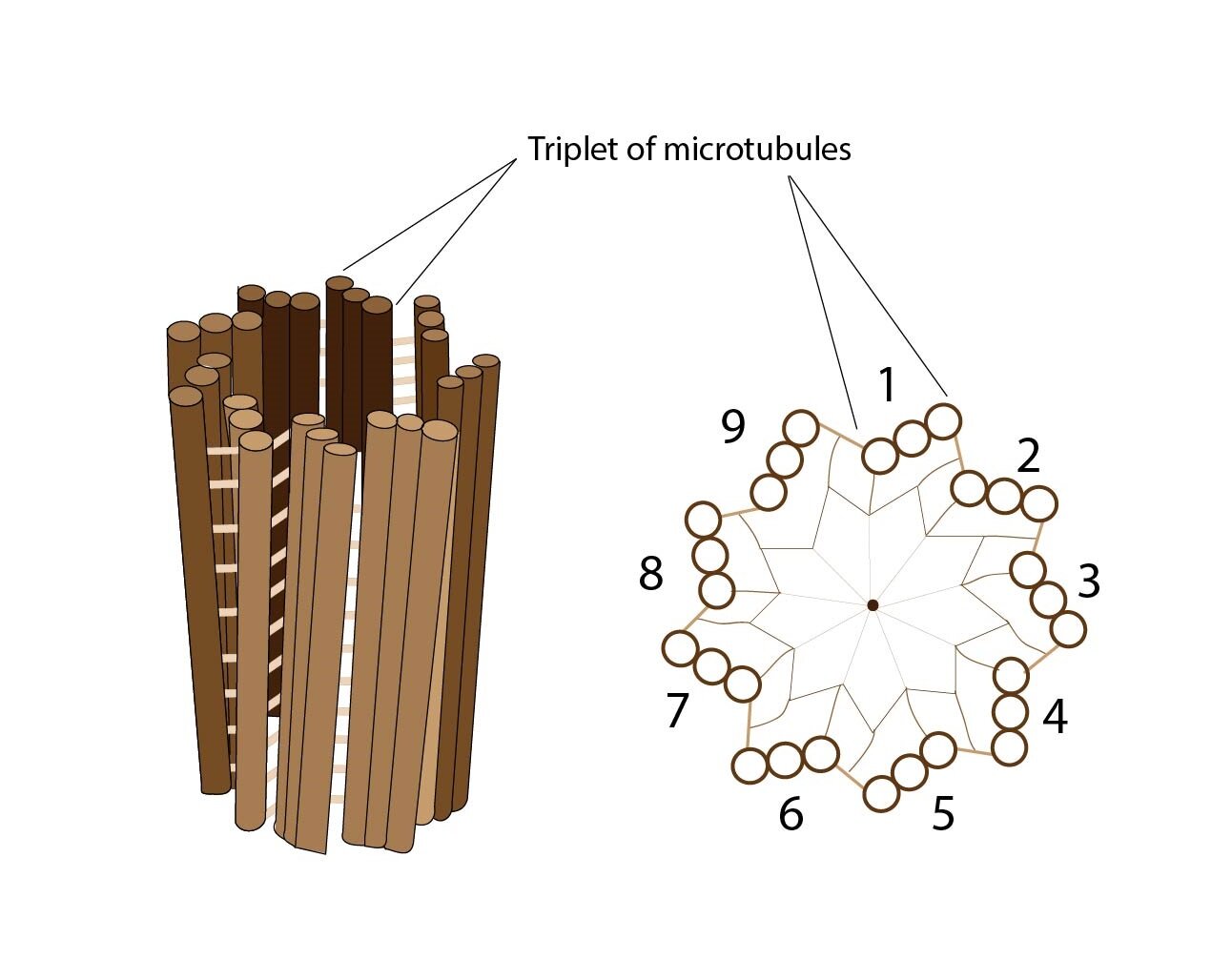
Another important cytoskeletal structure is centriole composed of typically arranged microtubules – 9 triplets of microtubules arranged parallely into the circle. In each cell are present two centrioles. During the cell division are located on poles of cell but in non-dividing cells, they are located near nucleus and Golgi apparatus.
Kinocilium is special motile structures located on the surface of pseudostratified columnar epithelium (e.g. respiratory epithelium). They are formed by circularly arranged 9 duplets of microtubules with two central microtubules. the axis of movement is along the central microtubules – the movement is faster in one direction and slower in another. The same structure is found in flagellum (except the length – flagellum is approximately 10x longer).
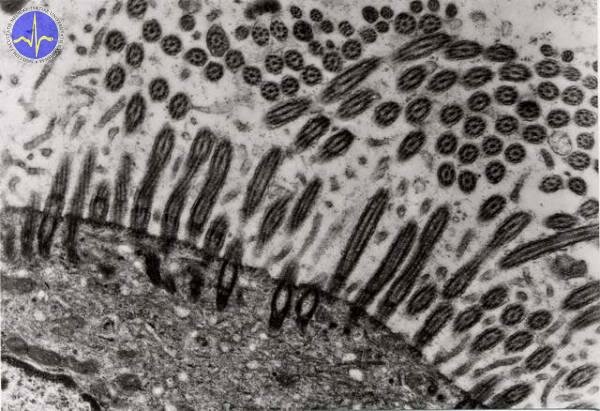
Kinocilia
Subchapter Author: Martina Šajdíková
![]()