Náplň podkapitoly:
1. Úvod do metabolismu proteinů a aminokyselin
2. Degradace aminokyselin
3. Degradace uhlíkatých skeletů aminokyselin
4. Tvorba neesenciálních aminokyselin v lidském těle
5. Významné deriváty jednotlivých aminokyselin
_
Úvod do metabolismu proteinů a aminokyselin
Proteiny patří mezi nejdůležitější a současně nejhojněji zastoupené biomolekuly lidského těla – celkově jich máme asi 14 kg (platí pro muže o průměrné váze 70 kg). Souhrn všech aminokyselin v těle nazýváme aminokyselinový pool. Jak se tento pool tvoří? A jak ho využíváme?
U zdravého dospělého člověka je asi 300-500 g proteinů denně degradováno na aminokyseliny, hovoříme o tzv. proteolýze. Další zdroj aminokyselin, proteiny obsažené v potravě, představují přibližně 70-100 g za den. Posledním zdrojem aminokyselinového poolu je biosyntéza neesenciálních aminokyselin tvořící denně 30-40 g aminokyselin. Přibližně stejné množství aminokyselin, jako uvolňuje proteolýza, se zpětně inkorporuje do proteinů, jedná se o proces tzv. proteosyntézy. Kolem 120 g aminokyselin denně degradujeme. V tomto procesu si můžeme řetězec aminokyseliny rozdělit na aminoskupinu (a ostatní atomy dusíku) a na uhlíkový řetězec – každý z nich má zcela odlišný osud, který si popíšeme dále v této podkapitole. Aminokyseliny mohou také sloužit jako prekurzory mnoha pro tělo významných látek – např. biogenních aminů, hemu či purinů a pyrimidinů.
Poločas života proteinů se výrazně liší protein od proteinu, a není tedy dost dobře možné uvést jeho průměrnou hodnotu. Obecně se ale dá říci, že strukturní proteiny jsou trvalejší (a mají tedy delší poločas), kdežto molekuly mnoha enzymů naopak existují jen velmi krátce velmi – jen několik desítek minut či hodin.
Proteolýza je kompletní degradace proteinů na volné aminokyseliny. V průběhu proteolýzy se uplatňují enzymy proteázy a peptidázy, které nacházíme jak v trávicím traktu, tak i v každé buňce – lyzosomy. Základní dělení peptidáz rozlišuje:
1) Exopeptidázy – amino- a karboxypeptidázy – štěpící proteiny / peptidy na koncích jejich řetězců
2) Endopeptidázy – trypsin, chymotrypsin či pepsin – štěpící vnitřní vazby proteinů / peptidů
Některé proteiny se degradují pomocí tzv. ubikvitin-proteazomové cesty. Ubikvitin, malý buněčný protein, se vyskytuje ve všech eukaryotních buňkách. Slouží jako značka, která předurčuje protein k jeho degradaci v proteazomech – proces nazýváme ubikvitinace (popřípadě polyubikvitinace – pokud je navázáno více molekul ubikvitinu).
Historická korelace:
Za objev rozkladu bílkovin iniciovaného ubikvitinem obdrželi v roce 2004 Nobelovu cenu za chemii („for the discovery of ubiquitin-mediated protein degradation”) Aaron Ciechanover, Avram Hershko a Irwin Rose.
Esenciální a postradatelné aminokyseliny
Pouze některé aminokyseliny (neesenciální, postradatelné) si naše tělo dovede samo nasyntetizovat. Zbylé (esenciální, nepostradatelné) musíme přijímat v potravě. Zde uvádíme jejich přehled:
Nepostradatelné (esenciální) aminokyseliny
1) Rozvětvené: Val, Leu, Ile
2) Aromatické: Phe, Trp
3) Bazické: Lys
4) Obsahující síru: Met
5) S hydroxylovou skupinou: Thr
Podmíněně esenciální aminokyseliny
Arg, His
Postradatelné aminokyseliny
Gly, Ala, Ser, Pro, Cys, Tyr, Asn, Gln, Asp, Glu
Důležité reakce aminokyselin
1) Dekarboxylace neboli odstranění karboxylové skupiny – vznikají biogenní aminy
2) Transaminace neboli výměna aminoskupiny s 2-oxokyselinou – vznikají 2-oxokyseliny
3) Oxidativní deaminace neboli oxidativní odstranění aminoskupiny – vznikají 2-oxokyseliny
4) Tvorba peptidové vazby – vznik peptidů a proteinů
Defekty metabolismu aminokyselin
U člověka existuje mnoho různých genetických defektů metabolismu aminokyselin. V případě mnoha z nich se v těle hromadí meziprodukty zapříčiňující defektní vývoj nervového systému, který často vyústí v mentální retardaci.
_
Degradace aminokyselin
Existuje 20 (21, pokud počítáme i selenocystein) základních proteinogenních aminokyselin, které mohou být v procesu translace vloženy do molekul proteinů. Katabolismus jejich uhlíkatých skeletů pokrývá přibližně 10-15 % energetických nároků našeho těla. Aminokyseliny také mohou sloužit jako substráty (prekurzory) pro biosyntézu ostatních živin – sacharidů (glukoneogeneze) a lipidů.
Nyní si probereme zvlášť osud aminodusíku a metabolismus uhlíkatého skeletu aminokyselin.
Odstranění aminoskupiny
Odstranění aminoskupiny vnímáme jako klíčový krok katabolismu aminokyselin. Dusík z aminoskupin se nedá využít pro produkci energie a musí být z našeho těla odstraněn. To se děje jednak jeho přeměnou na močovinu (asi z 95 %), následně vyloučenou z organismu močí, jednak jeho uvolněním v tubulárních buňkách ledvin z glutaminu jako NH3/NH4+ (asi z 5 %).
Transaminace aminokyselin
Transaminace jsou volně reverzibilní reakce katalyzované transaminázami (aminotransferázami). Během transaminace se vymění aminoskupina α-aminokyseliny s oxoskupinou 2-oxokyseliny – z aminokyseliny se vytváří 2-oxokyselina a z původní 2-oxokyseliny vzniká aminokyselina.
Aminoskupina je během reakce transportována ve vazbě na kofaktor pyridoxalfosfát (PLP, derivát vitaminu B6), který ji přenáší na oxokyselinu (tvorba Schiffovy báze).
Většina aminokyselin prochází při své degradaci transaminací. Jako konkrétní příklady transamináz si můžeme uvést aspartátaminotransferázu (AST) a alaninaminotransferázu (ALT), jež se běžně stanovují ke zjištění potenciálního poškození jaterních buněk. Jimi katalyzované reakce zachycuje následující schéma.
Vzniklé 2-oxokyseliny (oxaloacetát a pyruvát) se zapojují do energetického metabolismu buněk.
Existují ale i výjimky (např. threonin), jež se při své degradaci vůbec přímo netransaminuji
Přeměna glutamát / glutamin
Přeměnu karboxylové skupiny glutamátu (v postranním řetězci) na amidovou skupinu glutaminu katalyzuje cytosolický enzym glutaminsyntetáza. K reakci je kromě enzymu potřeba ATP a NH4+. Tato reakce slouží v buňkách CNS jako hlavní detoxikační mechanismus odstraňující toxický NH3 z mozkové tkáně. Vznikající glutamin je nejvýznamnější transportní forma aminodusíku (amoniaku) v krvi – zajišťuje transport z extrahepatálních tkání krví do jater a ledvin. Má nejvyšší plazmatickou koncentraci ze všech aminokyselin – 0,6 mmol/l (alanin – 0,3 mmol/l). V jeho molekule „skladujeme” dvě aminoskupiny/amoniaky. Glutamin současně dovede vnášet amoniak do různých biosyntéz – např. do tvorby purinových bází.
Uvolnění NH3 z glutaminu katalyzuje mitochondriální enzym glutamináza (hydrolytická deaminace, hojně se vyskytuje v hepatocytech a buňkách tubulů ledvin). Vzniklý amoniak se v jaterních mitochondriích zapojuje do močovinového cyklu, v ledvinách je vyloučen do moči, kde slouží jako její pufr.
Oxidativní deaminace
Během oxidativní deaminace se za souběžného uvolnění NH3 aminoskupina přeměňuje na ketoskupinu. Glutamát je jediná aminokyselina, která se v lidském těle deaminuje dostatečnou rychlostí. Přeměnu katalyzuje glutamátdehydrogenáza uložená v matrix mitochondrie (hlavně jaterních buněk). Tento proces demonstruje následující reakce:
Glutamát + NAD+ → α-ketoglutarát + NH4+ + NADH + H+
Vzniklý NH4+ vstupuje do močovinového cyklu a α-ketoglutarát se může využít v transaminacích či v Krebsově cyklu. Uvedená reakce je plně reverzibilní – z α-KG a NH4+ můžeme nasyntetizovat glutamát.
Závěrem této části tedy můžeme konstatovat, že většina aminokyselin prochází při své degradaci transaminací a že většina aminodusíku z aminokyselin se přímo či nepřímo nakonec koncentruje v molekule glutamátu/glutaminu. Z nich se následně uvolňuje v glutaminázové a glutamátdehydrogenázové reakci.
Močovinový (ornitinový) cyklus
Toxicita amoniaku
Amoniak volně prochází tělesnými bariérami, např. i hematoencefalickou bariérou. Při zvýšení jeho koncentrace v těle se vychýlí rovnováhy mnoha významných reakcí. Podívejme se na následující příklady:
Glutamát + NAD+ → α-ketoglutarát + NH4+
Glutamát + NH4+ + ATP → glutamin + ADP + Pi
Při nadbytku amoniaku se tedy pomalu zvyšuje koncentrace glutaminu, jehož tvorba ale současně spotřebovává α-ketoglutarát z Krebsova cyklu – postupně klesá rychlost této významné dráhy a tím i produkce energie v buňkách. Plazmatická koncentrace amoniaku by neměla překročit hodnotu 35 μmol/l. V lidském těle se většina toxického amoniaku přeměňuje reakcemi močovinového cyklu na močovinu.
Reakce močovinového cyklu
Urea, netoxická sloučenina, je transportována krevním řečištěm do ledvin, kde se z organismu vyloučí močí. Močovinový cyklus lokalizovaný jak v matrix mitochondrie, tak v cytosolu jaterních buněk představuje energeticky náročný proces, do něhož vstupují tři substráty – amoniak, oxid uhličitý (bikarbonát) a aspartát (jeho aminoskupina). Regulačním enzymem je mitochondriální karbamoylfosfátsyntetáza I. Ornitinový cyklus komunikuje s Krebsovým cyklem skrze oxalacetát a fumarát.
Tvorba močoviny probíhá během pěti reakcí:
1) Tvorba karbamoylfosfátu katalyzovaná mitochondriální karbamoylfosfátsyntetázou I:
NH4+ + HCO3– + ATP → karbamoylfosfát + 2 ADP + Pi
2) Tvorba citrulinu katalyzovaná ornitintranskarbamoylázou:
Ornitin + karbamoylfosfát → citrulin + Pi
Citrulin je transportován do cytosolu.
3) Tvorba argininsukcinátu katalyzovaná argininosukcinátsyntetázou:
Citrulin + Asp + ATP → argininsukcinát + AMP + PPi
4) Rozpad argininsukcinátu katalyzovaný argininosukcinázou:
Argininsukcinát → arginin + fumarát
5) Hydrolýza argininu katalyzovaná arginázou:
Arginin + H2O → ornitin + močovina
Následuje přenos ornitinu do mitochondriální matrix
Močovinový cyklus je těsně propojen s Krebsovým cyklem – ze vznikajícího fumarátu se stává aspartát. Jak tento vztah probíhá? Fumarát se nejdříve hydratuje na malát, jehož oxidací vzniká oxalacetát. Ten je prostřednictvím enzymu aspartátaminotransferázy transaminován s glutamátem, čímž vzniká aspartát vstupující do ornitinového cyklu. Glutamát se získává transaminací degradovaných aminokyselin, které předávají své aminoskupiny na molekuly α-ketoglutarátu.
Regulace ornitinového cyklu
Karbamoylfosfátsyntetáza I, hlavní regulační enzym ornitinového cyklu, je aktivována prostřednictvím N-acetylglutamátu, který vzniká reakcí AcCoA a glutamátu katalyzovanou N-acetylglutamátsyntetázou. Její aktivitu zvyšuje aminokyselina arginin. Transkripce enzymů močovinového cyklu se zvyšuje u vysokoproteinové diety či u narůstajícího proteokatabolismu (např. za hladovění), tedy ve stavech zvýšené nabídky aminokyselin. Protože močovinový cyklus patří mezi protonproduktivní reakce, nastává jeho útlum při poklesu pH – acidóze.
Glukóza-alaninový cyklus
Alanin se jednak spolupodílí na přenosu amoniaku krví, jednak slouží skrze pyruvát jako významný zdroj uhlíků pro proces glukoneogeneze – viz: Podkapitola 2/9. Glukóza-alaninový cyklus je meziorgánová metabolická dráha probíhající mezi svalovými buňkami a játry. Poté, co pyruvát vznikne ve svalových buňkách, podléhá transaminaci za vzniku alaninu. Ten se uvolní do krve, která jej transportuje do jater, kde se transaminací zpětně přeměňuje na pyruvát, jenž se může zapojit do procesu glukoneogeneze. Vzniklá glukóza se krví dostává do svalů a celý cyklus se uzavírá. Přenesená aminoskupina (amoniak) směřuje do močovinového cyklu.
_
Degradace uhlíkatých skeletů aminokyselin
V úvodu jsme uvedli, že se v těle vyskytuje 20 (21, pokud počítáme i selenocystein) proteinogenních aminokyselin. Různě složitá degradace jejich uhlíkatých skeletů obsahuje různý počet kroků. V tomto textu se nebudeme věnovat detailnímu průběhu všech katabolických drah, ale omezíme se jen na základní obecné mechanismy a několik vybraných příkladů.
Katabolismus uhlíkatých skeletů aminokyselin vyúsťuje v tvorbu sedmi produktů: pyruvát, acetyl-CoA, acetoacetyl-CoA, α-ketoglutarát, suc-CoA, fumarát a oxalacetát. Ty mají v energetickém metabolismu různý osud. Jde především o to, zda se z nich (a tudíž i z příslušných aminokyselin) mohou tvořit molekuly glukózy, či jen molekuly lipidů (mastné kyseliny a ketolátky). Podle toho rozlišujeme tzv. glukogenní a ketogenní aminokyseliny. Mezi ketogenní aminokyseliny patří ty, které vedou k tvorbě acetyl-CoA a acetoacetyl-CoA – Leu a Lys (začínají na písmeno L). Mezi glukogenní aminokyseiny řadíme ty, jež vedou k tvorbě zbylých pěti produktů – pyruvátu, α-ketoglutarátu, suc-CoA, fumarátu či oxalacetátu – serin, threonin, cystein, methionin, aspartát, glutamát, asparagin, glutamin, glycin, alanin, valin, prolin, histidin a arginin. Existují i aminokyseliny se dvěma degradačními produkty – jeden z nich je glukogenní a druhý ketogenní. Nazýváme je keto- i glukogenní aminokyseliny – patří mezi ně isoleucin, fenylalanin, tyrosin a tryptofan.
Následující přehled zachycuje, jaké aminokyseliny se degradují na jaké produkty:
1) Acetyl-CoA a acetoacetyl-CoA – čistě ketogenní jsou Lys a Leu, několik dalších aminokyselin poskytuje degradační produkty glukogenní i ketogenní – Phe, Tyr, Trp, Ile
2) α-ketoglutarát – pětiuhlíkaté aminokyseliny – Glu, Gln, Pro, Arg a His
3) Suc-CoA – nepolární aminokyseliny – Met, Ile a Val
4) Fumarát – Phe, Tyr
5) Oxalacetát – čtyřuhlíkaté aminokyseliny – Asp a Asn
6) Pyruvát – Cys, Ala, Ser, Gly, Thr, Trp
Degradace větvených aminokyselin – Val, Leu a Ile
Pro tyto aminokyseliny je příznačné, že jsou degradovány nikoli v jaterních buňkách, ale převážně v extrahepatálních tkáních – vysoká aktivita zejména ve svalových buňkách. Ty obsahují specifickou transaminázu produkující příslušné α-ketokyseliny – tzv. ketoanaloga větvených aminokyselin. Tato transamináza absentuje v jaterních buňkách. Ketoanaloga se přeměňují na deriváty acyl-CoA působením dehydrogenačního komplexu, který katalyzuje oxidativní dekarboxylaci a dehydrogenaci.
Genetický defekt tohoto dehydrogenačního komplexu vyvolává onemocnění nazývané maple syrup urine disease neboli nemoc javorového sirupu. Při tomto relativně vzácném onemocnění dochází k hromadění příslušných α-ketokyselin v tkáních a tělesných tekutinách (podmiňují charakteristický zápach moči po javorovém sirupu – páleném cukru). Defekt vyvolává abnormální vývoj mozku, mentální retardaci a může vyústit až v úmrtí jedince.
_
Tvorba neesenciálních aminokyselin v lidském těle
V lidském těle nemůžeme syntetizovat esenciální aminokyseliny – Phe, Trp, Val, Leu, Ile, Met, Thr a Lys. Dvě aminokyseliny jsou esenciální v období růstu organismu (období jejich zvýšené spotřeby), kdy rychlost jejich syntézy nestačí k pokrytí tělesných nároků – tzv. podmíněně esenciální aminokyseliny – Arg, His. Ostatní aminokyseliny řadíme mezi neesenciální aminokyseliny. Zde uvádíme přehled prekurzorů aminokyselin:
1) Oxalacetát → Asp, Asn
2) α-ketoglutarát → Glu, Gln, Pro, (Arg a His)
3) Pyruvát → Ala
4) 3-fosfoglycerát → Ser, Cys a Gly
5) Phe → Tyr
Fenylketonurie (PKU)
Fenylketonurie je autozomálně recesivní onemocnění (výskyt 8-10 případů / 100 000 jedinců) podmíněné absencí či sníženou aktivitou fenylalaninhydroxylázy. Ta fyziologicky katalyzuje hydroxylaci Phe na Tyr. Při defektu enzymu dochází k alternativnímu odbourávání Phe – vzniká fenylpyruvát (transaminace), fenyllaktát, fenylacetát či fenylethylamin. Tyto látky se hromadí v tkáních a tělesných tekutinách a vyvolávají typický zápach moči po myšině. Některé z nich způsobují závažné poškození mozku. Fenylketonurie byla prvním objeveným lidským genetickým defektem metabolismu aminokyselin a v současné době je jedním z onemocnění, u kterého se provádí screening u všech novorozenců. Pokud ji totiž dokážeme rozpoznat ještě v tomto věku, můžeme zabránit poškození mozku přísnou dietou s omezením Phe.
_
Významné deriváty jednotlivých aminokyselin
Dekarboxylace – tvorba biogenních aminů
Dekarboxylaci (odstranění karboxylové skupiny) podléhají jen některé aminokyseliny. Výsledkem je vznik tzv. biogenních aminů (monoaminů), které v lidském těle vykazují široké spektrum funkcí. Zde uvádíme jejich základní přehled:
1) Tyr → katecholaminy (DOPA →dopamin → noradrenalin → adrenalin)
2) Trp → serotonin (5-hydroxytryptamin)
3) Glu → γ-aminobutyrát (GABA)
4) His → histamin
5) Ser → etanolamin → cholin → acetylcholin
6) Cys → cysteamin
7) Asp → β-alanin
Oxid dusnatý (NO)
Oxid dusnatý je vazodilatační látka produkovaná endotelovými buňkami. Tvoří se z L-argininu reakcí katalyzovanou NO-syntázou:
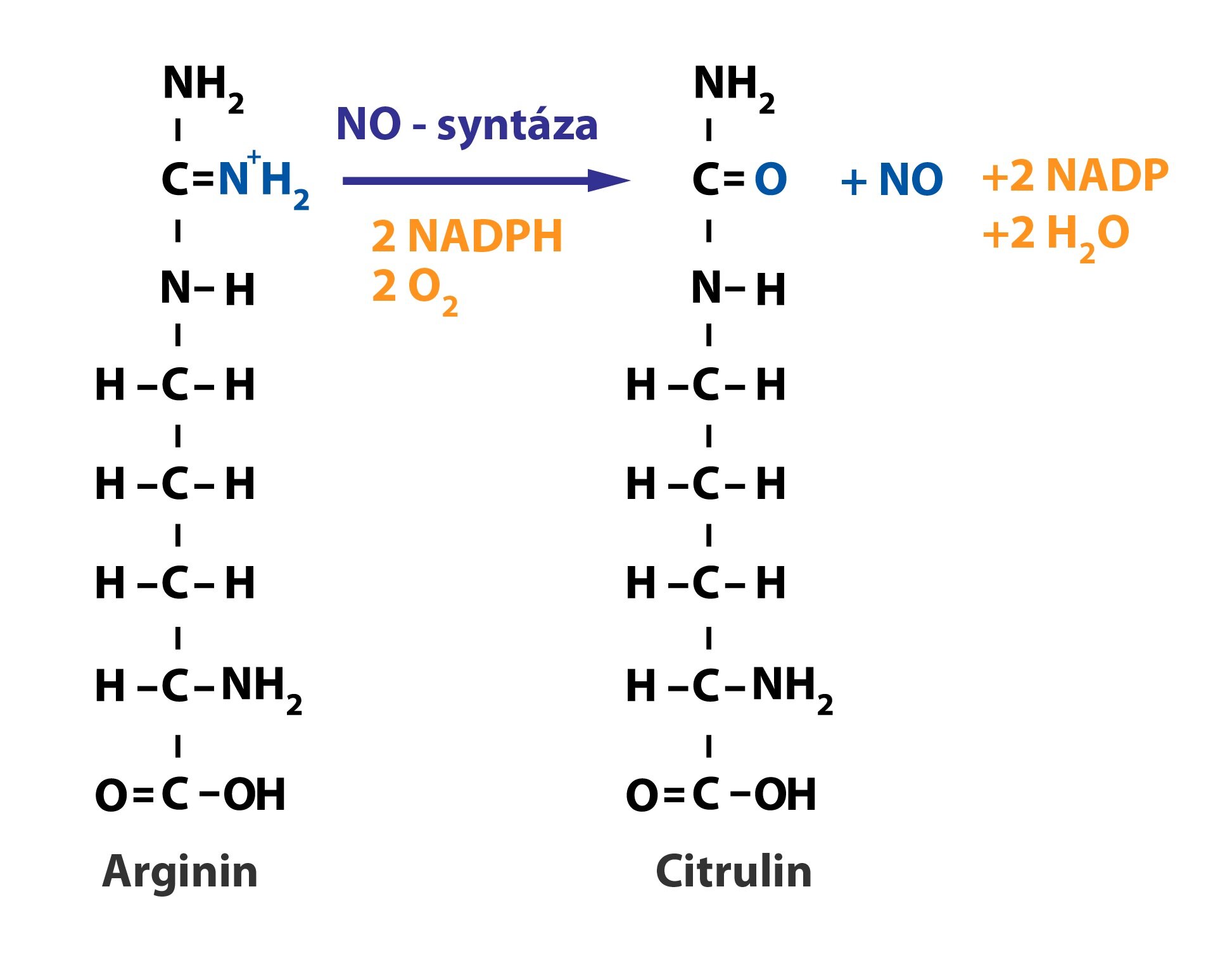
NO-syntáza se vyskytuje nejen v endotelových buňkách, ale i v některých buňkách imunitního systému (je jednou z cytotoxických látek) a v některých neuronech. O mechanismu a významu jeho působení podrobně pojednávají Kapitola 5 a Kapitola 10.
Další významné deriváty aminokyselin
Trp → melatonin
Phe a Tyr → hormony štítné žlázy, melanin
Gly → hem, puriny, kreatin, konjugace se žlučovými kyselinami
Arg a Ornitin → kreatin, polyaminy (spermidin, spermin)
Gly, Glu, Asp → puriny a pyrimidiny
Cys → taurin
_
Autoři podkapitoly: Josef Fontana and Petra Lavríková
![]()