Content:
1. Introduction to the degradation and the synthesis of glucose
2. Glycolysis
3. Gluconeogenesis
_
Introduction to the degradation and the synthesis of glucose
Saccharides are one of the main nutrients for heterotrophic organisms. Saccharides can be found in each cell of the human body. Saccharides have many different functions: (1) source of energy, (2) source of carbon atoms for syntheses, (3) reserves of energy (glycogen), (4) structural function (proteoglycans).
Glucose (Glc) is universal energetic substrate. One gram of glucose when oxidized provides 17 kJ (= 4 kcal). Of great importance is fact that our cells are capable of draining energy from glucose even in anaerobic conditions – this is not true for any other nutrient we use. There are some populations of cells that are strictly dependent on glucose – i.e. erythrocytes, cells of central nervous system, etc…. We would like to emphasize that pyruvate dehydrogenase reaction (PDH) is irreversible therefore it is not possible to synthesise glucose from fatty acids. Au contraire our cells are capable of converting excessive glucose into fatty acids and then into TAG.
Glucose blood concentration is called glycemia. Glycemia is normally 3,3 – 5,6 mmol/l, glycemia can after a meal increase to 7,0 mmol/l. In physiological conditions glucose is not present in the urine. In case that glycemia is higher than 10,0 mmol/l the renal threshold is exceeded and glucose gets into the urine. This condition is called glycosuria. Thus when glycemia is higher than renal threshold (value of threshold is 10,0 mmol/l) glucose can be found in the urine.
In the food is glucose in several forms: (1) free glucose, (2) part of oligosaccharides (predominantly disaccharides), and (3) part of polysaccharides. To the blood is from intestine released only free glucose.
Sources of blood glucose:
1) Saccharides from meal (digestion of saccharides in the GIT) or conversion of other nutrients from meal to glucose.
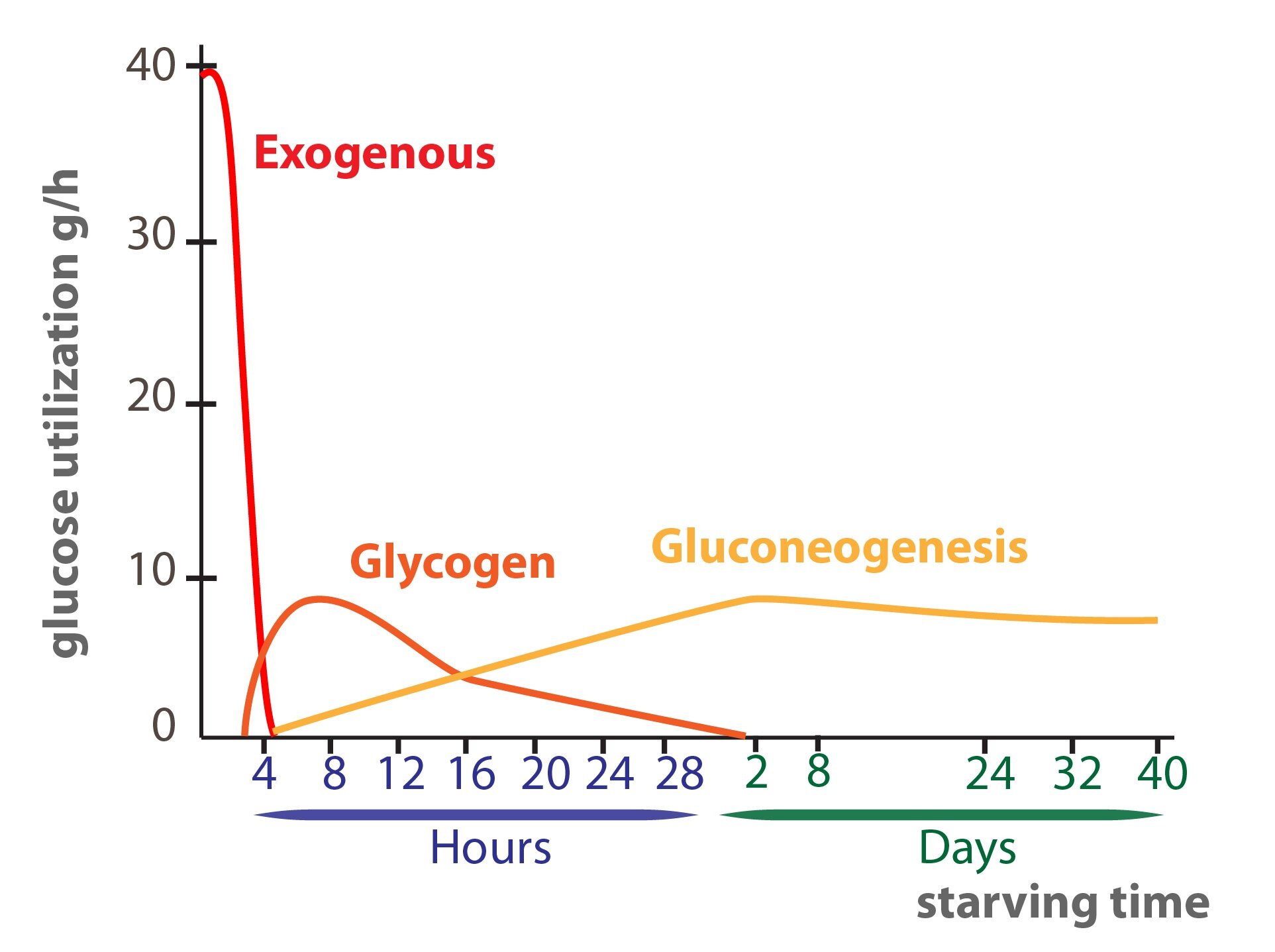
2) Degradation of liver glycogen. This source is predominantly important in the periods of time between meals. Supplies of glycogen are able to provide glucose for 24 hours.
3) Gluconeogenesis from C3 and C4 substances (lactate, glycerol, majority of amino acids). This source provides glucose in long-term fasting or in pathological conditions. In conditions of prolonged starvation glucose provides only 20% of energy used by body, rest is provided predominantly by lipid oxidation.
Acceptors of blood glucose
1) Tissues dependent on glucose (brain, erythrocytes) – independent on insulin
2) Tissues independent on glucose, thus able to use other substrates (e.g. skeletal muscles) – dependent on insulin
3) Glycogen synthesis in the liver, muscles and other tissues
4) Excessive glucose is converted to fatty acids and TAG. TAGs are stored in adipose tissue
5) Synthesis of many important substances (other monosaccharides, monosaccharide derivatives, etc…)
Saccharides are metabolized as phosphoric esters. Key compound of saccharides metabolism is glucose-6-phosphate (Glc-6-P). Glc-6-P connects many metabolic pathways: glycolysis, gluconeogenesis, the pentose pathway, synthesis of glycogen, glycogenolysis. Since Glc-6-P is not able to pass cytoplasmic membrane it keeps glucose in the cells. Glucose phosphorylation maintains concentration gradient between extracellular and intracellular space. This gradient eases entry of glucose to the cells.
Glycolysis is used by higher organisms as a main pathway for saccharide degradation. The pentose pathway is complementary pathway for saccharide degradation.
Mechanisms of glucose transport across cell membrane
Glucose is transported across cell membrane by two mechanisms: (1) facilitated diffusion (GLUT transporters) or (2) secondary active transport (SGLT transporters).
There are many glucose transporter in the cell membrane that work on the principle of facilitated diffusion. Facilitated diffusion is passive process. Glucose transfer is run by concentration gradient. The most important transporters are GLUT 1-7 (GLUcose Transporter). Only GLUT 4 is insulin-dependent. Insulin binds to its receptor and number of GLUT 4 transporters in the particular cell membrane is increased.
Summary of the basic GLUT transporters follows:
GLUT 1 and GLUT 3 are used for basal glucose uptake in tissues dependent on glucose (the brain, erythrocytes, the kidneys, and the placenta).
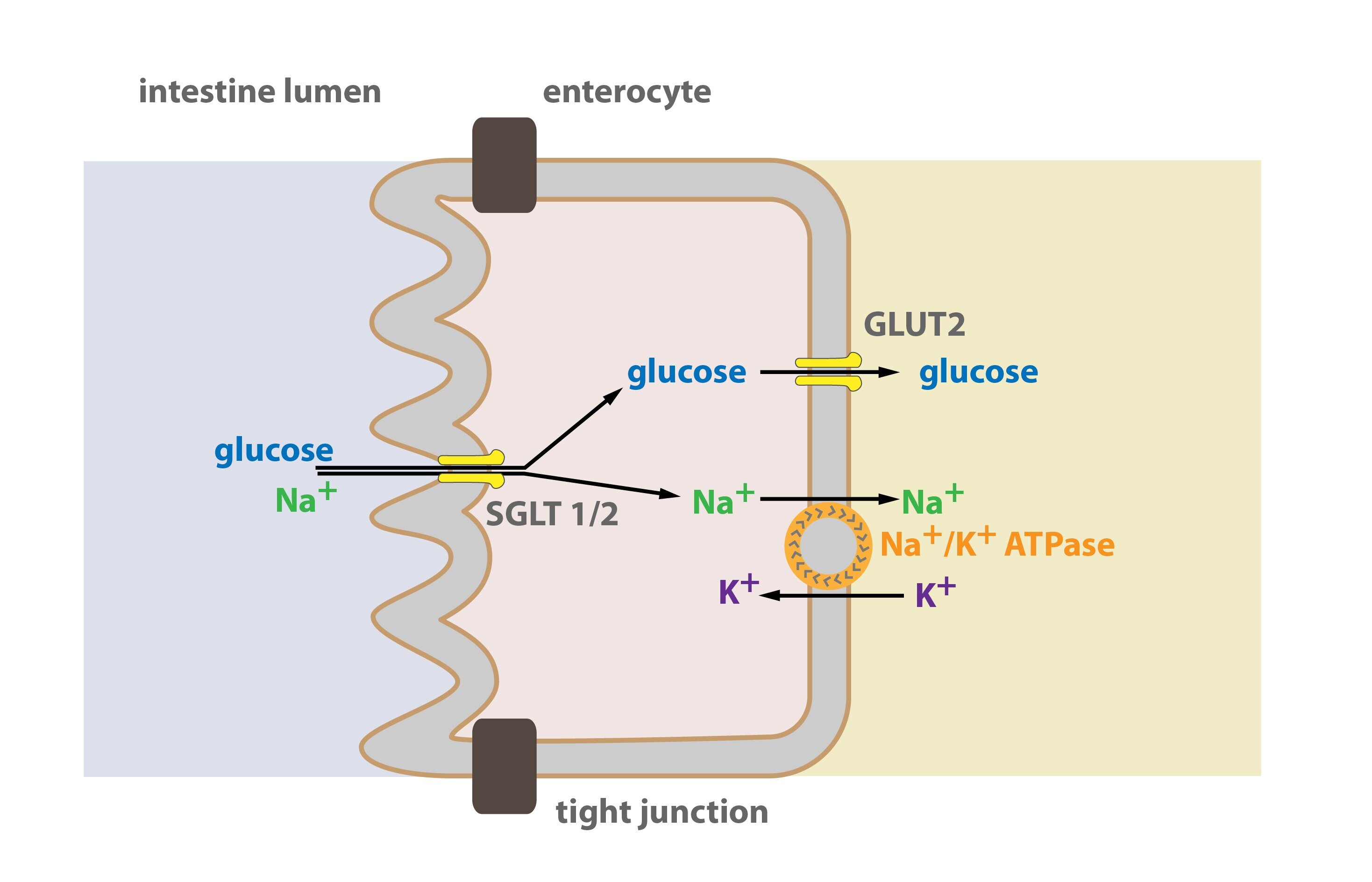
GLUT 2 is in the membranes of the pancreatic β-cells, hepatocytes and resorbing epithelia (proximal tubule of the nephron, intestine enterocytes). In hepatocytes and resorbing epithelia it allows transport of glucose from cells to the blood.
GLUT 4 is in the membranes of cells in glucose-independent tissues (i.e. in skeletal muscles, myocardium, and adipose tissue). GLUT 4 is incorporated to the membrane in conditions when insulin levels are elevated (i.e. in conditions of increased glycemia). This is natural after a meal. After a meal insulin-dependent tissues degrade 80 % of blood glucose. During fasting is glycemia low thus insulin is low, therefore GLUT 4 is not in the cell membranes and glucose is spared for glucose-dependent tissues. GLUT 4 is incorporated to the cell membrane from vesicles that are in the cytoplasm.
Active transport transports glucose in enterocytes and proximal tubule cells. Glucose is co-transported with Na+. Glucose is transported against its concentration gradient to the cell. This movement is driven by energy provided by Na+ ions that move to the cells according to their concentration gradient. This type of transport is provided by SGLT-1 and 2 (Sodium Glucose Transporter). Na+ is transported back to the ECF by Na+/K+ ATPase (this consumes ATP, hence it is active transport). Glucose is transported passively, Na+ provides energy from active transport, therefore glucose is transported so called secondary actively.
Glycolysis
Glycolysis (also Embden-Meyerhof-Parnas pathway) is basic metabolic pathway localized in cytoplasm of each cell in the body. Its nature is catabolic. One glucose molecule is converted to two three-carbon molecule – pyruvate (Pyr) (pyr can be converted to lactate (Lac)). Glycolysis has many functions, for example: (1) source of energy, (2) source of acetyl-CoA (AcCoA is substrate for lipid synthesis). Glycolysis can take place in both aerobic, and anaerobic conditions. In aerobic conditions glycolysis yields two molecules of pyruvate, two molecules of ATP and two molecules of NADH. In anaerobic conditions is pyruvate converted to lactate. This reaction spends two molecules of NADH. This however is advisable because two molecules of NAD+ are regenerated.
Glycolysis reactions
Overall equation of glycolysis is as follows:
Glucose + 2 NAD+ + 2 ADP + 2 Pi → 2 pyruvate + 2 NADH + 2 ATP + 2 H2O
Glycolysis can be divided into these phases:
1) investment of energy and activation of glucose
2) splitting of hexose to two trioses
3) trioses oxidation and energy production
4) conversion of pyruvate to lactate (in anaerobic conditions)
We will describe its single steps:
1) Glucose phosphorylation
Glucose is immediately phosphorylated when it enters the cell. This reaction converts neutral molecule of glucose to anion. Only this modified glucose (1) can be metabolised and (2) cannot pass across the membrane. Equation describes this step:
Glucose + ATP → Glc-6-P + ADP
This step is catalysed by the enzyme hexokinase, or glucokinase.
Glucokinase (= hexokinase type IV) is localised only in hepatocytes and in pancreatic β-cells. Hexokinase (not type IV) is in every tissue. These isoenzymes differ not only in localization, but also in physical-chemical characteristics. Glucokinase has high value of KM = 10 mmol/l. Glucokinase is thus activated in high glucose concentration. Therefore it acts predominantly after meals when glucose concentration in blood is high and it is needed to absorb glucose to the liver (e.g. for glycogen synthesis). Pancreatic β-cells in high glycemia increase insulin secretion. Hexokinase has KM = 0,1 mmol/l, hence it is fully active all the time (normal glycemia is 3,3-5,6 mmol/l and KM of hexokinase is 0,1 mmol/l). Hexokinase is regulated by another mechanism, it is negative feedback (i.e. it is inhibited by its own product – Glc-6-P). We can say that hexokinase produces as much Glc-6-P as the cells is able to utilize in its pathways. When Glc-6-P accumulates hexokinase is inhibited. Hexokinase is capable of phosphorylation of fructose.
2) Isomerisation of Glc-6-P to Fru-6-P
Glc-6-P is isomerised to Fru-6-P. This step is reversible and catalysed by the enzyme hexose phosphate isomerase.
3) Phosphorylation of Fru-6-P to Fru-1,6-BP utilizing ATP
Fru-6-P is phosphorylated to Fru-1,6-BP by the enzyme 6-phosphofructo-1-kinase. This enzyme is key allosteric regulatory enzyme of glycolysis.
In steps 1-3 two molecules of ATP per one molecule of glucose were invested.
4) Aldol cleavage of Fru-1,6-BP to two phosphorylated trioses
Fru-1,6-BP is cleaved to two phosphorylated trioses – glyceraldehyde-3-P (Gra-3-P, aldose) and dihydroxyacetone phosphate (DHA-3-P, ketose). This step is catalysed by the enzyme aldolase. Aldolase is lyase. There are two isoenzymes (aldolase A and B).
5) Trioses isomerisation
Glyceraldehyde-3-P and dihydroxyacetone-3-phosphate can be converted to each other by the enzyme triosephosphate isomerase. This reaction is of great importance because only glyceraldehyde-3-P is able to continue in glycolysis, hence this step continually replenish its cytosolic pool.
6) Glyceraldehyde-3-P oxidation to 1,3-bisphosphoglycerate
This reaction is only oxidation in whole glycolysis. This step is catalysed by the enzyme glyceraldehyde-3-phosphate dehydrogenase. This reaction yields: (1) reduced cofactor NADH and (2) 1,3-bisphosphoglycerate (1,3-BPG). 1,3-BPG contains great amounts of energy. Reaction is exergonic – i.e. oxidation produces new COO– group and Pi is bound to it with macroergic anhydride bond.
7) Conversion of 1,3-bisphosphoglycerate to 3-phosphoglycerate
1,3-BPG is hydrolysed to 3-phosphoglycerate (3-PG). This step is catalysed by the enzyme phosphoglycerate kinase. This reaction is one of the substrate phosphorylations – i.e. ADP is converted to ATP.
8) Isomerisation of 3-phosphoglycerate to 2-phosphoglycerate
This step is catalysed by the enzyme phosphoglycerate mutase.
9) 2-phosphoglycerate dehydration to phosphoenolpyruvate (PEP)
Dehydration of 2-phosphoglycerate is catalysed by the enzyme enolase. This step yields macroergic compound – phosphoenolpyruvate. In PEP phosphate group is ester bound.
10) Phosphoenolpyruvate conversion to pyruvate
At first Pi is cleaved then unstable enol-pyruvate is produced. Enol-pyruvate is isomerised to more stable keto-pyruvate. This conversion releases great amounts of free energy, i.e. this step is strongly exergonic and practically irreversible. Released energy is used for synthesis of ATP from ADP – substrate phosphorylation. This step is catalysed by the regulatory enzyme pyruvate kinase.
In steps 4-10 two molecules of ATP per one three carbon fragment (pyruvate) are produced. Energetic balance is +2 moles of ATP per 1 mole of glucose (2 ATP utilized, 4 ATP produced).
Metabolic fates of pyruvate
Pyruvate is branching point of glycolysis. Fate of pyruvate depends on oxidative state of the cell – NADH must be reoxidised to NAD+.
In aerobic conditions pyruvate is transported to mitochondrial matrix. In mitochondria is pyruvate converted to acetyl-CoA by the enzyme pyruvate dehydrogenase. Acetyl-CoA can enter the TCA cycle. Reduced cofactor NADH is not able to pass across the mitochondrial membrane (membrane is impermeable for whole molecule). NADH is therefore used for reductions of some compounds in cytosol: e.g. oxaloacetate to malate or dihydroxyacetone phosphate to glycerol-3-P. These products are able to get through mitochondrial membrane and transport reducing equivalents to the mitochondrial matrix. This mechanism is denoted as shuttle mechanism (or just shuttle). There are two shuttles: (1) glycerol-phosphate and (2) malate-aspartate. Above mentioned reactions in mitochondria take place in inverse course:
Malate + NAD+ → oxaloacetate + NADH
Glycerol-3-P + FAD → dihydroxyacetone-phosphate + FADH2
(here different cofactor accepts reducing equivalents)
Reduced cofactors enter the ETC, where their regeneration and oxidative phosphorylation take place (thus NAD+ and ATP are produced). OAA returns to cytosol but not directly. OAA requires transamination to aspartate that is catalysed by the enzyme aspartate aminotransferase (AST). In cytosol inverse reaction take place (i.e. OAA is produced).
In anaerobic conditions (e.g. intensely working muscle with diminished blood supply) or in erythrocytes lactate is produced from pyruvate. Lactate is released to the blood stream. NAD+ is regenerated. Conversion of pyruvate to lactate is catalysed by the enzyme lactatedehydrogenase (LDH):
Pyruvate + NADH + H+ → lactate + NAD+
NAD+ produced by this reaction is co-enzyme for glyceraldehyde-3-phosphate dehydrogenase that is essential for glycolysis. Produced lactate can (1) enter Cori cycle (see below), (2) be oxidised in tissues with aerobic metabolism (heart, liver) to CO2 and H2O. Accumulating lactate causes decreased pH that leads to the muscle pain and fatigue.
You should notice now that aerobic glycolysis produces much more ATP than anaerobic glycolysis.
2,3-BPG shunt
2,3-BPG (2,3-bisphosphoglycerate) shunt is diversion from glycolysis in erythrocytes. 1,3-BPG is converted to 2,3-BPG. 2,3-BPG does not contain macroergic phosphate thus when converted to 3-PG ATP is not produced, just Pi is released. This means that erythrocyte gains less ATP in glycolysis. 2,3-BPG production is however very important because it is necessary for regulation of haemoglobin affinity to oxygen, thus it regulates oxygen transport in the blood.
Glycolysis regulation
There are 3 regulatory steps in glycolysis: (1) 6-phosphofructo-1-kinase, (2) pyruvate kinase, (3) hexokinase. These enzymes catalyse irreversible exergonic reactions.
6-phosphofructo-1-kinase (PFK-1)
Phosphofructokinase is allosteric enzyme regulated by several activators and inhibitors. It is the main glycolysis regulatory step:
1) Increased ATP/AMP ratio lead to glycolysis inhibition
Glycolysis is pathway that leads to ATP production. ATP is both substrate and allosteric inhibitor of this enzyme. AMP is activator of this enzyme. In excess of ATP glycolysis is inhibited thus glucose is not utilized as nutrient.
2) Citrate inhibits glycolysis
In case that fatty acids are oxidised acetyl-CoA is produced. AcCoA inhibits PDH. Thus pyruvate is carboxylated to OAA. In situation when there is enough AcCoA, citrate is produced. Citrate accumulates in front of isocitrate dehydrogenase and leaks to the cytoplasm. In cytoplasm citrate indicates that there are enough substrates for the TCA cycle and thus it inhibits regulatory steps of glycolysis.
3) Fructose-2,6-bisphosphate (Fru-2,6-BP)
Fru-2,6-BP activates glycolysis. Insulin increases its concentration. In case that insulin:glucagon ratio is increased 6-phosphofructo-2-kinase (PFK-2) is activated and Fru-2,6-BP is produced.
4) Glycolysis is activated by insulin, inhibited by counterregulatory hormones
Increased insulin:glucagon ratio lowers intracellular cAMP. This leads to superiority of dephosphorylation processes. Decreased ratio insulin:glucagon or increased counterregulatory hormones lead to superiority of phosphorylation processes. PFK-2 is active when dephosphorylated.
5) Acidic pH inhibition
PFK-1 is inhibited by protons. Both pyruvate and lactate are quite strong acids and their significant accumulation could lead to damage of the cell. Because glycolysis is important source of these acids their increased concentration (and thus lowered pH) leads to inhibition of the regulatory enzyme.
Hexokinase and pyruvate kinase are of lesser importance. Hexokinase is both inhibited by its product (i.e. Glc-6-P) and activated by insulin. Pyruvate kinase is regulated by covalent modification regulated by insulin:glucagon ratio.
Clinical correlation:
Relatively common enzyme defects is congenital deficiency of pyruvate kinase. Erythrocytes are affected mostly because less ATP is produced. Thus they are not capable of (1) maintaining their membrane integrity (active membrane transports) and (2) their shape. This results in increased haemolysis. Haemolytic anaemia develops.
In hypoxia (insufficient oxygenation of the tissues) lactate is produced in increased amounts. Lactate is quite strong acid and causes acidification of the body. This is called lactate acidosis.
_
Gluconeogenesis
Gluconeogenesis is process of glucose production from non-saccharide compounds. Used precursors are predominantly three- and four-carbon compounds – e.g. lactate, pyruvate, glycerol, Ala, Gln and other glucogenic amino acids and propionate (especially in ruminants). Gluconeogenesis is localized in both mitochondrial matrix, and cytosol, predominantly in hepatocytes, tubular cells of the kidneys and in enterocytes. We are able to survive prolonged starving thanks to gluconeogenesis, because glycogen stores are depleted in 24 hours of fasting. Gluconeogenesis is however slightly activated even every morning after night fasting.
Gluconeogenesis proceeds like glycolysis but in reverse course. You should now recall that some of glycolysis reactions are irreversible and therefore in gluconeogenesis it is necessary to use different enzymes. Since there are 3 such enzymes these steps are denoted as bypasses 1, 2 and 3. In glycolysis are irreversible steps catalysed by the enzymes: (1) pyruvate kinase, (2) 6-phosphofructo-1-kinase, (3) hexokinase/glucokinase.
Gluconeogenesis reactions
Now we will describe single reactions of gluconeogenesis:
1-2) Bypass 1 – conversion of pyruvate to phosphoenolpyruvate
First step is transport of pyruvate to the mitochondrial matrix. Pyruvate is carboxylated to oxaloacetate. This reaction is catalysed by the enzyme pyruvate carboxylase, ATP is utilized, and cofactor is carboxybiotin. Oxaloacetate is transported from mitochondria to cytosol (transamination to Asp or reduction to malate). In cytosol is OAA regenerated and converted to phosphoenolpyruvate. This conversion is catalysed by the enzyme phosphoenolpyruvate carboxykinase (utilizes GTP).
3-8) Reactions 3 to 8 are in essence reverse reactions of glycolysis and are localised in cytosol
9) Bypass 2 – conversion of Fru-1,6-BP to Fru-6-P
Fru-1,6-bisphosphatase hydrolyses Fru-1,6-BP to Fru-6-P. This reaction is key regulatory step of gluconeogenesis.
10) Fru-6-P isomerisation to Glc-6-P
11) Bypass 3 – conversion of Glc-6-P to free glucose
Glucose-6-phosphatase hydrolyses Glc-6-P to free glucose (i.e. catalyses cleavage of phosphate). This enzyme is localised in smooth endoplasmic reticulum. Glc-6-P is transported to the endoplasmic reticulum by the enzyme translocase. This separation to the endoplasmic reticulum has important reason – i.e. it is not desirable to get free glucose phosphorylated to Glc-6-P. Free glucose is released to the blood stream.
Energetic balance of gluconeogenesis
Gluconeogenesis is energetically demanding process. One molecule of glucose synthesis spends 6 macroergic phosphates. Overall equation follows:
2 Pyr + 4 ATP + 2 GTP + 2 NADH + 4 H2O → Glc + 4 ADP + 2 GDP + 6 Pi + 2 NAD+ + 2 H+
Substrates for gluconeogenesis
Lactate
Lactate is one of the main sources of carbon atoms for gluconeogenesis. Lactate is produced in anaerobic glycolysis from pyruvate by the enzyme lactatedehydrogenase (LDH). Main producers of lactate are working muscle cells and erythrocytes. Lactate is released to the blood stream, transported to the liver and converted to glucose. Glucose is released to the blood and muscle cells, erythrocytes can utilize this glucose. This cycle is called the Cori cycle.
Pyruvate
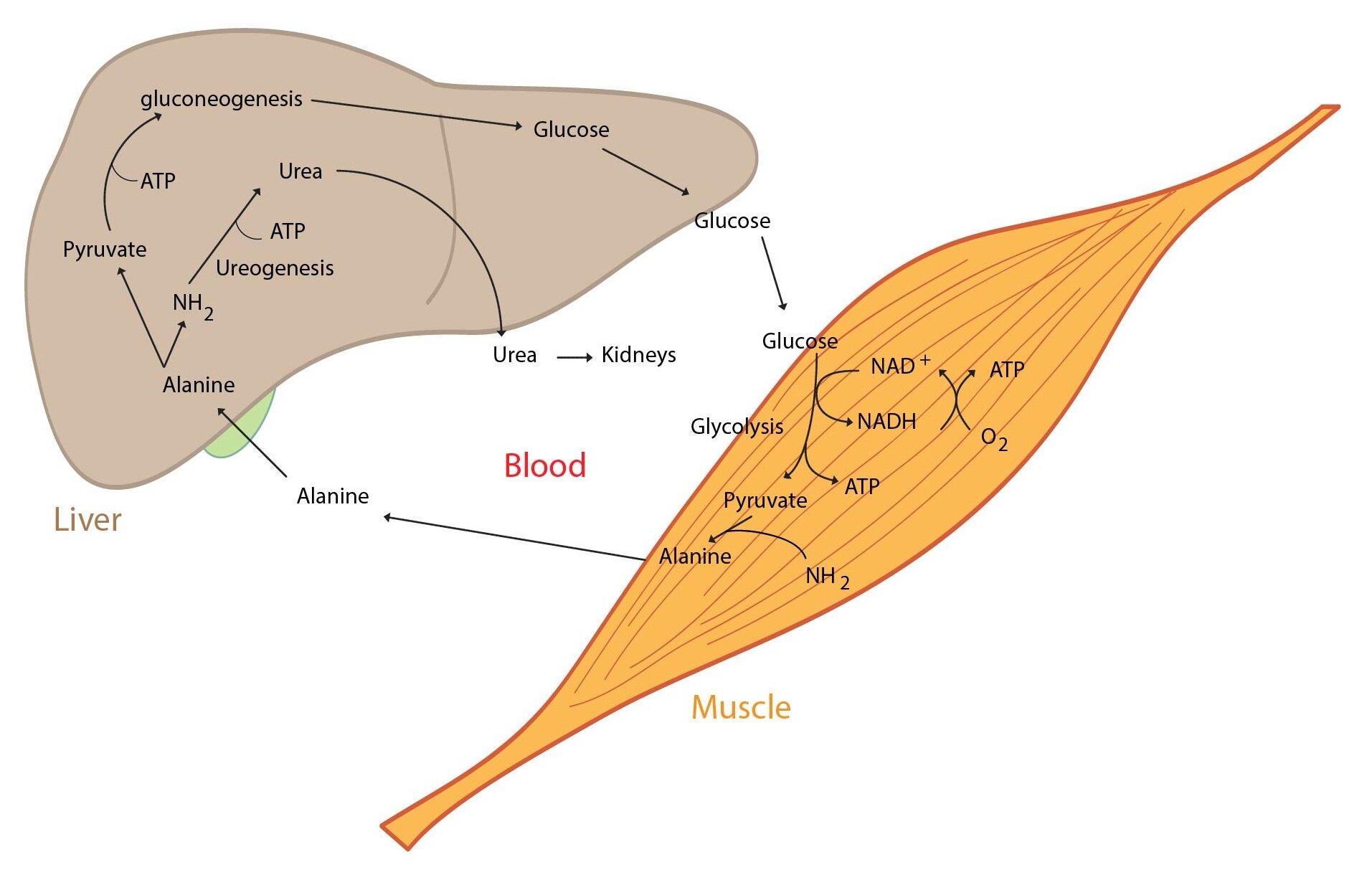
Pyruvate is produced in many peripheral tissues. Description of the glucose-alanine cycle now follows. This cycle is performed between muscle cells and liver. Pyruvate is produced in muscle cells, transaminated to alanine. Alanine enters blood and is transported to the liver. Alanine is in the liver converted to pyruvate that enters gluconeogenesis. This yields glucose that enters blood and is utilised by muscle. Thus cycle encloses.
Glucogenic amino acids
Carbon skeletons of all amino acids except leucine and lysine are source of carbon atoms for gluconeogenesis. The most important are alanine and glutamine. Detailed mechanism is beyond the aim of this text. The most important source of glucogenic amino acids are muscle proteins.
Glycerol
Glycerol is produced in triacylglycerol hydrolysis. At first glycerol is phosphorylated to glycerol-3-P. This step is catalysed by the enzyme glycerol kinase. Then glycerol-3-P is dehydrogenated to dihydroxyacetone-phosphate by the enzyme glyceraldehyde-3-phosphate dehydrogenase. Dihydroxyacetone-P is an intermediate of gluconeogenesis.
Sources of energy
Energy for gluconeogenesis is obtained from β-oxidation of fatty acids. In starving fatty acids are released from store triacylglycerol of adipose tissue and are metabolised in the liver.
Regulation of gluconeogenesis
Gluconeogenesis is metabolic pathway that is activated in starving or in pathological conditions (stress as a result of infection, polytrauma etc…)
Regulatory steps of gluconeogenesis are those that bypass irreversible reactions of glycolysis:
1) Pyruvate carboxylase: activated by acetyl-CoA (e.g. from β-oxidation of fatty acid)
2) PEP carboxykinase, Fru-1,6-bisphosphatase, Glc-6-phosphatase: regulated by the very same factors as glycolysis but in reverse course. Fru-1,6-bisphosphatase is activated by citrate, inhibited by AMP or Fru-2,6-BP
Next important factor influencing gluconeogenesis is supply of substrates (substrate are produced by proteolysis, lipolysis).
Insulin inhibits gluconeogenesis, counterregulatory hormones (glucocorticoids, glucagon, catecholamines) stimulate gluconeogenesis.
_
Subchapter Author: Josef Fontana a Petra Lavríková
![]()